
Схема и техника наращивания ресниц лисий эффект
Чтобы подчеркнуть глубину взгляда, девушки используют подводки и туши для ресниц с эффектом увеличения. Однако, если природные реснички тонкие и маленькие, ни какая тушь не позволит сделать их объемными и выразительными. В этом случае, не помощь придет процедура наращивания, которую предлагают в каждом салоне красоты. Особо популярностью пользуется технология наращивания с эффектом лисий взгляд.
Лисий эффектОб особенностях методики
Искусственное увеличение объема и численности ресниц предполагает приклеивание к каждой природной волосинке одной или двух синтетических. Для фиксации ворсинок используют прозрачный или черный клей, создающий эффект макияжа. В качестве материала для наращивания применяют синтетические ворсинки, напоминающие шелк, соболиный или норковый мех. Мастера не рекомендуют использовать натуральные ворсинки, поскольку это может вызвать аллергию.
Эффект от наращивания ресниц держится от 4 до 8 недель, при условии прохождения коррекции каждые 14-20 дней.
О разных эффектах
Мастера, занимающиеся искусственным увеличением объема и численности ресниц, знают, что методика позволяет добиться нескольких эффектов:
- Натуральный.
- Кукольный.
- Эффект взгляда белки.
- Лисий взгляд.
Каждый эффект подходит к определенному типу лица, разрезу глаз. Лисий взгляд подходит девушкам округлой формой глаз на фото. Благодаря наклеиванию длинных ворсинок на внешней стороне век, получается визуально удлинить разрез и поменять форму очей.
Секреты технологии
Лисий эффект наращивания ресниц на схеме на фото, выглядит, как плавное увеличение объема и численности волосинок от внутреннего угла глаз к внешнему. Задача косметолога обеспечить плавность перехода.
Наращивание ресниц 2д с лисьим эффектом, может осуществляться методом поресничного или пучкового увеличения. В первом случае, каждая природная волосинка увеличивается двумя искусственными. Во втором – пучок из двух-пяти ворсинок сразу приклеивается к одной реснице. Отзывы утверждают, что первый вариант позволяет получить более природное обрамление глаз.
Характеристики ресниц
Кроме выбора методики, длины и количества искусственных ворсинок, важно подобрать правильный цвет. Традиционно наращивание осуществляют волосинками черного и коричневого цвета. Но возможные и другие, яркие варианты. Для создания праздничного образа, используют зеленые, синие, красные и многоцветные волосинки. Для особых случаев, реснички могут быть украшены стразами, блестками.
Немаловажным параметром, который следует учесть при выборе эффекта и схемы наращивания, является степень изогнутости ворсинок. Она может быть слабой или значительной. Если природное обрамление век редкое, лучше применять 3d эффект, позволяющий в три раза увеличить объем и количество волосинок вокруг глаз. Отзывы утверждают, что классическое 3d увеличение способно преобразить взгляд девушки.
Технология в деталях
Кому подходит лисий эффект наращивания ресниц? Девушкам, у которых редкие, небольшой длины реснички. Итак, чтобы преобразить природное обрамление век, необходимо очистить кожу от косметики и жирных природных выделений. Делать это нужно с помощью специальной обезжиривающей жидкости.
Следующий шаг – защита нижних век от попадания клея с помощью специальных силиконовых накладок. Благодаря силикону излишняя влага на ресничках испарится, что позволит волосинкам не слипаться.
Классическое наращивание с эффектом лисьего взгляда продолжается непосредственным приклеиванием искусственных ворсинок к каждой природной волосинке. Делать это нужно по схеме, очень аккуратно, чтобы клей и место фиксации не были заметны окружающим.
Делать это нужно по схеме, очень аккуратно, чтобы клей и место фиксации не были заметны окружающим.
Лисий эффект наращивания ресниц на фото, завершается расчёсыванием ресничек и необходимой коррекцией.
Рекомендации по уходу
- Первые 10-16 часов после процедуры не умываться, не тереть и не чесать глаза.
- Для сохранности эффекта наращивания по технике лисий эффект не посещать баню, сауну и другие места с повышенной влажностью.
- Отдыхать на спине, не касаясь 2д ресничками подушки.
- Расчесывать наращенные волосинки специальной щеточкой.
Кому подходит наращивание ресниц с лисьим эффектом на видео, расскажет опытный косметолог.
В завершении
Обладательницы округлых глаз, могут эффектно изменить природную форму путем наращивания искусственных ресниц по технике лисьего взгляда. Схема наращивания предполагает плавное увеличение численности и объема ворсинок от внутреннего угла к внешнему. Длинные, красивые реснички на внешней стороне век преображают взгляд, делая похожим его на лисий. Данный эффект можно добиться методом увеличения численности волосинок по 2д или 3d технике.
Схема наращивания предполагает плавное увеличение численности и объема ворсинок от внутреннего угла к внешнему. Длинные, красивые реснички на внешней стороне век преображают взгляд, делая похожим его на лисий. Данный эффект можно добиться методом увеличения численности волосинок по 2д или 3d технике.
Нарощенные ресницы лисий эффект. Лисий эффект наращивания ресниц: как его сделать правильно
Наращивание ресниц сейчас является одной из самых популярных косметических процедур среди представительниц прекрасного пола.С помощью опытного мастера, который подберет идеальную форму, длину, густоту и даже тип ресничек, можно визуально преобразить не только форму глаз, но и даже слегка изменить черты лица, добиваясь желаемой степени выразительности.
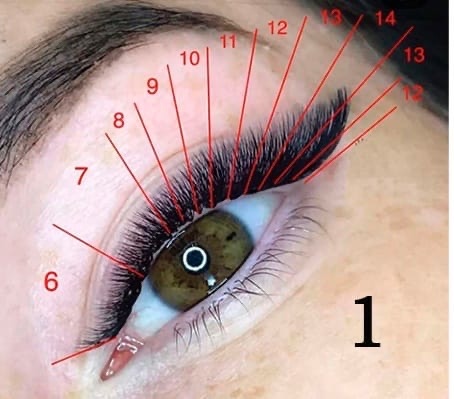
Эффект следует выбирать в зависимости от того, какую форму имеют ваши глазки. Например, обладательницам округлых глаз прекрасно подойдет лисий эффект наращивания (фото №1), о котором далее и пойдет речь.
Лисий эффект наращивания зрительно удлиняет глазки, как будто «вытягивая» их внешние уголки и образуя «стрелку».
Мастер приклеивает реснички разной длины, начиная от самых коротких у внутреннего уголка глаза и заканчивая длинными у внешнего. Этот процесс наглядно показан на схеме (фото №2).
В результате взгляд приобретает загадочность, глубину и красивую искорку хитрости, придающую женщине неповторимый шарм. Длина и плавность переходов подбирается индивидуально.
Если вы желаете, чтобы результат выглядел естественно, выбирайте реснички длиной 6-10 мм, если же хотите добиться более впечатляющего, но несколько ненатурального эффекта, можете смело останавливаться и на 14-15 миллиметрах.
Но исход процедуры наращивания зависит не только от длины и густоты ресничек, но и от их типа. Для наращивания используются:
- шелковые
- норковые
- соболиные
- колонковые ресницы.
Сейчас искусственные реснички для наращивания изготавливаются исключительно из высококачественных синтетических материалов, поскольку натуральные способны вызывать аллергию.
Все их названия условны, так как обозначают не материал, а скорее толщину и структуру. Например, «норковые» ресницы выглядят на глазах достаточно естественно, а шелковые идеальны для достижения объемного, яркого результата, причем их не понадобится дополнительно подкрашивать тушью.
Именно эти два типа чаще всего используются мастерами для создания безупречного лисьего эффекта.
Важный совет от редакции!
Все пользуются косметическими средствами, но исследования показали страшные результаты. Ужасная цифра года – в 97.5% популярных шампуней находятся вещества отравляющие наш организм. Проверьте состав вашего шампуня на наличие sodium lauryl sulfate, содиум лаурет сульфат, кокосульфат, ПЭГ. Эти химические вещества разрушают структуру локонов, волосы становятся ломкими, теряют упругость и силу.
Самое страшное то, что эта гадость через поры и кровь накапливается в органах и может вызывать онкологические заболевания. Мы настоятельно рекомендуем отказаться от использования средств, в которых находятся данные вещества.
Единственный производитель полностью натуральной косметики. Вся продукция производятся под строгим контролем систем сертификации. Рекомендуем к посещению официальный интернет магазин (mulsan.ru). Если сомневаетесь в натуральности вашей косметики, проверьте срок годности, он не должен превышать 11 месяцев.
Как проходит процесс наращивания?
Наращивание ресничек и последующее создание «лисьего» взгляда происходит в несколько этапов.
- Мастер очищает и обезжиривает сами ресницы и кожу вокруг глаз с помощью специальных косметических средств.
Накладываются коллагеновые подушечки, предотвращающие склеивание верхнего и нижнего ресничного ряда.
- После бережного подсушивания мастер начинает ювелирную процедуру наращивания (фото №3). Отделяя каждую природную ресницу, он приклеивает к ней искусственную, не затрагивая при этом слишком молодых, слабых и полупрозрачных ресничек (фото №4).
 Места склейки обязательно должны быть крепкими, микроскопическими и незаметными для невооруженного глаза. Это тщательно проверяется путем расчесывания и рассматривания.
Места склейки обязательно должны быть крепкими, микроскопическими и незаметными для невооруженного глаза. Это тщательно проверяется путем расчесывания и рассматривания. - После окончания процедуры коллагеновые подушечки снимаются. Готово!
 Места склейки обязательно должны быть крепкими, микроскопическими и незаметными для невооруженного глаза. Это тщательно проверяется путем расчесывания и рассматривания.
Места склейки обязательно должны быть крепкими, микроскопическими и незаметными для невооруженного глаза. Это тщательно проверяется путем расчесывания и рассматривания.Если вы желаете, чтобы процедура прошла комфортно, а результат выглядел по-настоящему красиво, отнеситесь к выбору мастера очень серьезно.
Профессионал не только подберет для вас оптимальную форму, длину и густоту ресниц, но и создаст желаемый лисий эффект, который будет идеально адаптирован для вашей индивидуальной формы глаз – округлые глазки зрительно удлинятся, а опущенные уголки поднимутся.
Чтобы максимально долго сохранять лисью «хитринку», ухаживайте за наращенными ресницами. Не наносите ежедневно обильный макияж, старайтесь не тереть глаза и не спать, уткнувшись в подушку личиком.
Каждая девушка имеет уникальную внешность, но не все умеют ее подчеркивать. Поэтому, процедура наращивания ресниц сейчас так популярна. В этой статье мы подробно рассмотрим лисий эффект наращивания ресниц фото и видео материалы нам в этом помогут.
В этой статье мы подробно рассмотрим лисий эффект наращивания ресниц фото и видео материалы нам в этом помогут.
Выбор ресниц
С помощью лисьего эффекта, глаз удлиняется, и ресницы образовывают стрелку. Это все благодаря тому, что с внутренних уголков глаз наклеиваются короткие реснички, и по пути к внешнему уголку – ресницы удлиняются. Длина ресничек подбирается индивидуально, исходя из формы лица и размера глаз.
С помощью наращивания можно изменить даже форму глаз. Лисий эффект подойдет лучше всего девушкам с круглыми глазами, лучше не делать такую процедуру обладательницам узких глаз.
Материалы должны быть высококачественными, чтобы не вызывали аллергию. Для наращивания используют такой материал:
- Норка.
- Шелк.
- Соболь.
Эти названия условные, ведь они означают не материал, а структуру волосков. Например, шелковые реснички будут очень выразительными и объемными, с ними вы забудете про тушь. Наиболее часто для данного вида наращивания используют норковые и шелковые ресницы.
Процесс
Лучше обратиться к дипломированному специалисту, ведь доверять свое здоровье кому-нибудь не следует, во избежание плачевных последствий. Чтобы выглядеть как лисичка, нужно пройти несколько этапов:
Обезжиривание
Мастер должен очистить кожу вокруг глаз и сами реснички, только после этого можно переходить к следующим пунктам. Для того чтобы ресницы не слипались, клеится скотч или ложится мягкая подушечка, которая разделяет верхний и нижний ряды век.
Наращивание
После того, как кожа чистая и сухая можно приступать к наращиванию. Мастер бережно отделяет каждую ресничку и клеит на нее накладной волосок. При этом слабые и короткие волоски мастер не трогает.
Если наклеить новую ресничку на маленький волосок, то в процессе роста она может изменить свое положение, что будет выглядеть не очень красиво. Слабые реснички тоже лучше не трогать, ведь со временем она может выпасть.
Процедура проходит медленно, чтобы новые реснички хорошо держались, и не было следов клея. Также, в процессе наращивания мастер прочесывает ресницы, чтобы не пропустить никакую ресничку.
Также, в процессе наращивания мастер прочесывает ресницы, чтобы не пропустить никакую ресничку.
Переход от коротких ресничек к длинным ресницам должен быть очень мягким и плавным, чтобы это выглядело максимально естественно. Чтобы взгляд был максимально изящным, нужно использовать как можно больше волосков разной длины. Принято использовать волоски до 10 мм, но если девушка очень яркая и любит привлекать внимание, то можно использовать и длинные волоски – до 16 мм.
Снятие подушечек
Когда все ресницы приклеены, мастер снимает подушечки, и вы можете любоваться своим выразительным и эффектным взглядом.
Коррекция
К сожалению, ничто не вечно и наращенные ресницы не исключение. В среднем, наращенные ресницы держатся 3-5 недель, все зависит от ваших индивидуальных способностей.
Свои ресницы отрастают, поэтому после 3-5 недель нужно делать коррекцию наращенных ресниц или снимать их и клеить новые.
Если у вас выпало не много ресниц, то целесообразнее для вас будет процедура коррекции. Мастер просто приклеит ресницы в промежутки и у вас снова будет выразительный и красивый взгляд.
Правила
Те, кто наращивают ресницы впервые, должны запомнить такие правила:
- найдите квалифицированного мастера;
- после процедуры наращивания нужно следить и ухаживать за ресницами;
- нельзя тереть глаза, чтобы не травмировать глаз и не повредить реснички;
- не злоупотребляйте макияжем;
- спите на боку или на спине, чтобы ресницы не терлись о подушку и не повредились из-за нее;
Правила не очень трудные, поэтому их легко придерживаться. Если будете следовать правилам. То ваши реснички продержаться максимально долго.
Противопоказания
К сожалению, не всем можно наращивать реснички таким способом. От лисьего эффекта лучше отказаться, если:
- у вас жирная кожа;
- проблемы с глазами, заболевания;
- носите контактные линзы. Ведь жидкость, в которой хранятся линзы, пагубно влияет на клей, убирая его клейкие способности, из-за чего реснички будут держаться недолго.
- аллергия на материал (на клей или на сами реснички).
 Ведь жидкость, в которой хранятся линзы, пагубно влияет на клей, убирая его клейкие способности, из-за чего реснички будут держаться недолго.
Ведь жидкость, в которой хранятся линзы, пагубно влияет на клей, убирая его клейкие способности, из-за чего реснички будут держаться недолго.Отнеситесь очень серьезно к выбору мастера, лучше, если у него будет много опыта. Ведь именно от этого зависит длительность процедуры (начинающий мастер может наращивать ресницы 5-6 часов), комфорт во время наращивания и самое главное – результат наращивания. Ведь с новыми ресничками вы должны восхитительно выглядеть.
Только профессионал может подобрать для вас нужную форму, длину, материал. Ведь опытный мастер сможет придать вашим глазкам необходимый лисий эффект, сделает правильную густоту, которая подойдет под вашу форму лица. Глаза зрительно удлинятся и уголки глаз соответственно приподнимутся.
В процессе носки не ленитесь ухаживать за ресницами.
Видео по теме статьи:
- обезжирить и очистить ресницы;
- для того чтобы не склеились верхние и нижние реснички, необходимо под глаза наложить специальный скотч, сейчас часто используют коллагеновые подушечки;
- подсушить реснички, чтобы они стали сухими, иначе плохо будут приклеиваться;
- к каждой живой ресничке приклеивается искусственная. Очень важно не трогать самые маленькие и слабые.
ВАЖНО ЗНАТЬ!
Не вздумайте покупать средства для РОСТА РЕСНИЦ, не прочитав отзывы по ним. ..
..
Красота — страшная сила! Каждая девушка хочет нравиться парням и поэтому всегда стремится выглядеть красивой и сексуальной. Наращивание ресниц Лисий взгляд поможет женскому полу привлечь на себя внимание. Действительно, первое, на что мужчины обращают внимание, — это глаза. Глазами можно заинтересовать, влюбить в себя, покорить и в то же время отпугнуть.
1 Кому подходит данный вид наращивания
Глаза — зеркало души. Как бы странно это ни звучало, но это истина. Поэтому за глазами нужен особый уход. Нельзя обходить стороной и ресницы. Сейчас очень распространенной является такая модная процедура, как . Способов наращивания очень много. Остановимся на одном из них — наращивание ресниц Лисий эффект.
Не всех женщин природа одарила естественными шикарными ресничками. С помощью наращивания профессиональный мастер сможет даже изменить , сделать взгляд широко распахнутым, сексуально узким, кошачьим, обеспечить Лисий эффект, приподнять глаза, подарить нежность.
Лисий эффект наиболее подойдет для обладательниц круглой формы глаз. Не подходит девушкам с узкими глазами. Запомните, выбирая тот или иной эффект наращивания ресниц, обратите внимание на себя, потому что у девушек с различными формами глаз один и тот же эффект будет смотреться по-разному. Очень важно делать эту процедуру у дипломированного мастера, а не у самоучки.
Данный вид наращивания визуально удлиняет глаза, делая взгляд необычным и заманчивым.
Приклеивая реснички, мастер плавно переходит от коротких к более длинным. Профессионализм мастера проявляется в том, насколько плавный переход он делает от коротких ресниц к ресницам побольше. От внутренних уголков глаз к внешним может быть от 2 до 3 видов длины. Чем больше ресничек разной длины задействовано, тем красивее взгляд. Чтобы взгляд был естественнее, используют реснички от 6 мм (внутренний уголок глаза) до 10 мм (внешний уголок глаза). Смелые обладательницы данного эффекта могут экспериментировать с более длинными — от 10 мм до 16 мм.
Кроме длины и густоты, на общий вид глаз также влияет материал, из которого сделаны реснички. Сегодня мастера используют чаще всего . Именно они наиболее подходят для данного наращивания.
2 Основные правила процедуры
Для того чтобы сделать наращивание с лисьим эффектом, нужно:
Если насаживать ресницы на маленькие, то, когда они вырастут, искусственная может изменить свое первоначальное закрепление (может перевернуться в противоположную сторону, «лечь» на соседние, торчать, портя общий вид). Слабые тоже желательно обходить стороной, потому что они в ближайшее время выпадут или прогнутся под тяжестью искусственной.
Основные правила для тех, кто впервые будет :
- Очень внимательно отнеситесь к выбору профессионального мастера. От него зависит, как будут выглядеть ваши глазки.
- Если вы уже сделали данную процедуру, то не забывайте о ежедневном уходе. Во-первых, не трите глаза. Во-вторых, минимум макияжа каждый день. В-третьих, не спите лицом в подушку. Следуя этим правилам, вы будете носить реснички долго. Это так практично и красиво! К сожалению, не все могут сделать себе данную процедуру. Кому противопоказано:
И немного о секретах…
История одной из наших читательниц JulieSem:
«Особенно удручали меня ресницы, потому что глаза у меня крупные, а вот ресницы стали короткие и начали постоянно выпадать. Я Часто пользовалась недорогими тушами, закручивала реснички щипчиками. Я долгое время не знала как решить проблему. Даже хорошая тушь не решала проблему. Как вернуть густые и ресницы и сделать их красивыми? А ведь ничто так не старит или молодит человека, как его глаза.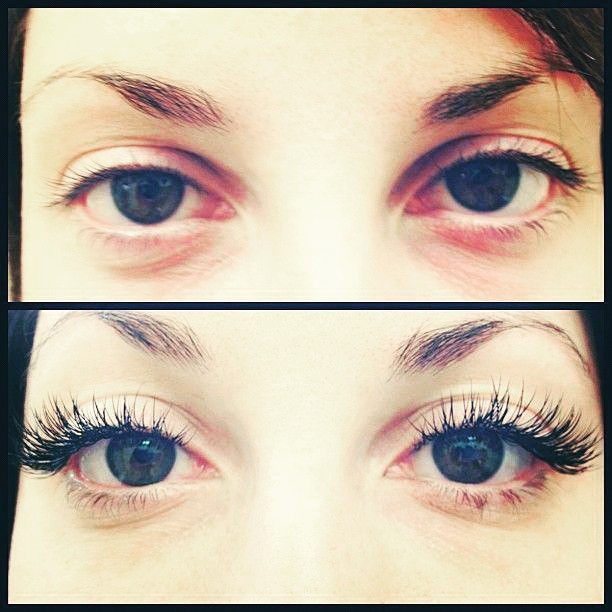
В общем, к 22 годам я перепробовала все методы, но вернуть былую красоту моим РЕСНИЦАМ, мне реально помог только один способ…»
Процедура наращивания ресниц очень популярна и востребована. К услугам мастеров обращаются молодые девушки и женщины в возрасте. Здесь нет возрастных границ и серьезных противопоказания, а конечный результат может серьезно изменить внешность, подарить молодость глазам, вернуть им свежий вид, глубину и налет сексуальности. Но часто можно услышать негативные отзывы об опыте наращивания. Это не всегда связано с профессиональными качествами лешмейкера и материалами, чаще женщины просто не соблюдают банальные правила по уходу за искусственным объемом. Даже банальная подводка нарощенных ресниц должна проводиться по правилам.
Наращивание искусственных волосков на глаза было придумано с целью оградить женщину от ежедневной необходимости проводить время перед зеркалом, накладывая макияж при помощи декоративной косметики. Это прекрасная альтернатива туши. Она никогда не течет от дождя и жары и не осыпается. Но не всем клиенткам достаточно выразительного объема, часто свою красоту им хочется подчеркнуть дополнительно при помощи подводки.
Она никогда не течет от дождя и жары и не осыпается. Но не всем клиенткам достаточно выразительного объема, часто свою красоту им хочется подчеркнуть дополнительно при помощи подводки.
Сочетается ли наращивание ресниц и макияж декоративной косметикой? Если дама всегда ярко красилась, то отказаться от этой привычки только из-за ресниц ей будет трудно. Да и часто в нашей жизни случаются такие моменты, когда нужно навести полноценный макияж на лице для праздника или фото. Конечно, никто не запрещает вам пользоваться тушью, подводкой, тенями, но делать это нужно аккуратно.
Основные правила подбора косметики для сочетания с нарощенными ресницами:
- Не рисуйте стрелку тушью и косметическим карандашом. Тушь имеет рассыпчатую структуру, будет стягивать кожу и портить внешний вид. Карандаш предполагает механическое воздействие, что при неосторожном применении может вызвать выпадение искусственных волосков. Лучше пользоваться специальными подводками или тенями.
- Нельзя пользоваться водостойкой косметикой, так как для ее удаления необходимо использовать специальные средства и требуются оказывать механическое воздействие руками.
- Тщательно подбирать краску, чтобы в ее составе не содержались жиры и масла, разрушающие сцепляющие свойства клея.

Специалисты не советуют накладывать макияж ежедневно, так как регулярные касания к искусственным ресницам способны вызвать их выпадение и повлиять на их внешний вид.
Как пользоваться подводкой с нарощенными ресницами
Если возникла необходимость в наложении стрелочки на верхнее веко, то проявите осторожность. Как уже было сказано, используйте для этой цели специальную подводку на водяной основе и с мягкой кисточкой, чтобы не оказывать механическое воздействие на глаза и не растворить клей.
Как делать подводку:
- Очистите кожу тоником или мицеллярной водой.
- Слегка припудрите или наложите тени.
- Аккуратно выньте кисточку из тюбика, чтобы на ней было достаточное количество краски и отсутствовали излишки.
- Прикройте глаз и начинайте вести линию по направлению от внутреннего уголка к внешнему. Движения должны быть уверенными и четкими.
- При допущении неровностей используйте ватную палочку, смоченную в воде.
- Если подводка попала на ресницы, то их также достаточно промыть мокрой ваткой. После высыхания они вернуться к прежнему виду.
 Движения должны быть уверенными и четкими.
Движения должны быть уверенными и четкими.Чтобы макияж держался долго и не оказывал негативного влияния на наращивание ресниц, пользуйтесь только качественной косметикой от известных брендов.
Так как в них используются натуральные компоненты, и состав указан на упаковке. Дешевая продукция часто содержит вредные вещества, о которых производитель предпочитает умалчивать.
Особенности техник наращивания с эффектом подводки
Часто на фото можно увидеть, как модели пленят роскошным объемом ресниц, который дополнительно подчеркнут затемнением у корней. В большинстве случаев этот эффект достигается в процессе наращивания, а не посредством создания стрелки косметикой.
Как создается эффект подводки? Существует три вариант достижения подобного результата.
- Использование черного клея-смолы для фиксации искусственных ресниц. Благодаря микроскопическим темным капелькам клеящего состава на основании роста волосков появляется визуальный результат подкрашивания.
- Наращивание по специальной технологии «Совершенная линия» или Perfect Line. методика предполагает наложение нескольких рядов искусственных волосков. Это похоже на объемное наращивание, но первый искусственный ряд делится на два и на них накладываются волоски разной длины – на верхний ряд длинные, на нижний – короткие или наоборот. В результате достигается двойной результат – искусственный объем служит на пару недель дольше и межресничное пространство становится более выразительным, появляются стрелки.
- Наращивание ресниц с эффектом Лисий взгляд. В этом варианте волоски крепятся от внутреннего уголка, к внешнему с равномерным увеличением длины, что создает видимость подводки.
Как видите, нет необходимости красить нарощенные ресницы, придавая им больше глубины и яркости, достаточно обратиться к хорошему мастеру, который подберет правильную технику наращивания.
Как смывать подводку
Если вы обладательница искусственного обрамления глаз и приходиться периодически краситься декоративной косметикой, то вы должны знать, как правильно смывать краску, не нанося вред ресницам.
- Для удаления косметики используйте средства для демакияжа без содержания жирных компонентов.
- Пользуйтесь ватной палочкой или диском.
- Не нужно тереть глаза руками.
Чтобы удалить стрелку, смочите ватный диск или палочку в воде или лосьоне и аккуратно смойте краску, не касаясь линии сцепления искусственного материала с натуральными ресницами. Только после полного очищения можно умыть лицо.
Нарощенные ресницы с эффектом стрелочки на веке выглядят намного выразительнее и ярче, чем обычный объем. Если хотите добиться именно этого эффекта, то подберите самый удобный способ. Для праздничных случаев и редких вечеринок вполне подойдет подводка, но если желаете ежедневно носить стрелочки на глазах, то лучше выберите наращивание с эффектом подведения века. Но прежде чем обращаться к лешмейкеру, изучите свои фото и отражение в зеркале, может пару недель применения средства для роста ресниц подарят желаемый эффект густоты и тогда можно будет без опаски пользоваться любимой косметикой.
Но прежде чем обращаться к лешмейкеру, изучите свои фото и отражение в зеркале, может пару недель применения средства для роста ресниц подарят желаемый эффект густоты и тогда можно будет без опаски пользоваться любимой косметикой.
По секрету…
- Вы – одна из тех миллионов женщин, которые хотят, чтобы их ресницы стали длиннее и гуще?
- А после наращивания ресниц состояние родных безумно ухудшилось?
- И вы уже задумывались о радикальных мерах?
Оно и понятно, ведь ресницы — это ваша красота и повод для гордости. Кроме того, это как минимум сейчас в моде. А то, что женщина, с привлекательными ресницами выглядит моложе – аксиома не требующая доказательств.
Существует много способов изменения своей внешности, среди которых особое место занимает наращивание ресниц. С помощью такой процедуры можно придать взгляду загадочность и выразительность, а также немного подправить редкие от природы ресницы и добавить им густоту и объем. Например, лисий эффект наращивания ресниц часто используют для визуального изменения формы глаз, в макияже артистов, а также при создании вечернего образа.
Во времена, когда эта процедура только появилась в индустрии красоты, она была достаточно дорогостоящей, и позволить себе сделать ее могли не все желающие, однако сегодня такое наращивание уже доступно каждой девушке. Существует несколько способов наращивания, которые создают разный эффект. Одним из них и является лисий эффект.
Подходит для девушек, имеющих круглую форму глаз, выпуклые глаза или приподнятые внешние уголки. Не рекомендуется выполнять такой способ наращивания девушкам, имеющим опущенные уголки глаз, так как это придаст дополнительную унылость внешнему облику. При маленьком разрезе глаз также не рекомендуется делать наращивание в таком стиле, так как глаза удлиняются и становятся еще меньше.
Данный эффект достигается путем приклеивания искусственных ресниц на все верхнее веко, при этом длина ресниц, расположенных во внешнем углу глаз, будет значительно длиннее, чем у внутреннего угла.
Увеличение длины должно быть плавным. При таком способе форма глаз становится более вытянутой и тем самым достигается нужная выразительность и дополнительный шарм. Длина ресниц подбирается мастером индивидуально для каждой девушки, в зависимости от состояния ее натуральных ресниц, а также от желаемого эффекта. Для получения более естественного эффекта следует выбирать длину искусственных ресниц до десяти миллиметров. Если нужен ошеломляющий эффект, то можно воспользоваться длиной до пятнадцати миллиметров.
Длина ресниц подбирается мастером индивидуально для каждой девушки, в зависимости от состояния ее натуральных ресниц, а также от желаемого эффекта. Для получения более естественного эффекта следует выбирать длину искусственных ресниц до десяти миллиметров. Если нужен ошеломляющий эффект, то можно воспользоваться длиной до пятнадцати миллиметров.
Ожидаемый результат можно получить только при тщательном подходе к выбору специалиста. Именно от работы мастера зависит возможность получить красивые и густые реснички и взгляд, как у лисички. Опытный косметолог всегда подберет нужную длину, а также подходящий качественный материал для наращивания, который, в свою очередь, избавит клиента от негативных последствий.
При выборе натуральных и качественных рабочих материалов достигается отсутствие у клиента аллергических реакций. Из всего многообразия представленных производителями материалов, для данного вида подходят соболиные и шелковые реснички, так как они обладают нужной длиной и толщиной. А при использовании последних не требуется дополнительного их подкрашивания тушью.
А при использовании последних не требуется дополнительного их подкрашивания тушью.
А теперь можно ознакомиться с процессом, как сделать лисий взгляд поэтапно.
Для этого понадобятся следующие инструменты: искусственные ресницы в необходимом количестве, пинцет, клей (черный или прозрачный), обезжириватель, специальные подушки для наращивания и щеточка. Перед самой процедурой специалист обрабатывает ресницы и поверхность века специальным средством, удаляющим с поверхности жир. Далее необходимо наклеить на нижнее веко по краю подушку, которая не будет позволять нижним ресницам мешать и склеиваться с верхними.
Затем необходимо к каждой реснице приклеить дополнительную, отодвигая остальные. Приклеивать следует обязательно к натуральным ресницам, отступая немного от линии роста. Расстояние от основания ресниц до места приклеивания должно быть не более 0,5 миллиметров. Если расстояние больше, то свободный край нарощенной ресницы будет причинять значительный дискомфорт. Наклеивать на новые растущие ресницы не рекомендуется, так как они только начинают свою фазу роста и еще слабы.
Наклеивать на новые растущие ресницы не рекомендуется, так как они только начинают свою фазу роста и еще слабы.
Прикреплять ресницы к коже века не следует, так как они долго не продержатся и отклеятся. Для крепления следует лучше использовать прозрачный клей, так как он не так заметен для окружающих. Черным клеем могут пользоваться профессионалы, которые имеют большой опыт наращивания, так как линия от клея должна быть ровной и напоминать линию подводки.
Каждая последующая ресница приклеивается только после полного высыхания предыдущей. Начиная с середины верхнего века, следует начинать увеличивать длину ресниц для создания нужного эффекта.
После того, как все реснички прикреплены, следует дождаться, пока они высохнут, и убрать подушку с нижнего века. Полученный результат можно посмотреть на фото.
После данной процедуры следует избегать косметики, содержащей в своем составе масла, так как они способствуют расщеплению клея и быстрой потере ресниц. Не рекомендуется спать лицом в подушку, чтобы реснички не замялись и не отклеились. Ежедневно следует их расчесывать специальной щеточкой.
Не рекомендуется спать лицом в подушку, чтобы реснички не замялись и не отклеились. Ежедневно следует их расчесывать специальной щеточкой.
Для выполнения наращивания ресниц есть и противопоказания.
Не стоит делать эту процедуру людям с повышенной слезоточивостью и чувствительностью глаз, а также при наличии аллергии на компоненты, используемые при наращивании.
В связи с тем, что жизнь ресниц коротка и длится около девяноста дней, то следует периодически делать коррекцию, чтобы поддерживать нужный эффект. Через полгода следует снять искусственные и восстановить свои ресницы с помощью специальных средств. Через пару месяцев можно выполнить наращивание снова.
Видео по теме статьи
Наращивание ресниц — Студия красоты и здоровья «Сирень»
Цена: от 2000 до 2500 р.
Длительность процедуры: 3 — 4 часа.
С помощью наращенных ресниц вы сможете стать обладательницей самых красивых, длинных и пушистых ресниц. Теперь уже Вам не нужно будет тратить время на ежедневные подкрашивания ресниц тушью и прочими средствами, и беспокоиться об их внешнем виде. По крайней мере, на протяжении месяца и даже более Вы сможете наслаждаться своим бесподобным внешним видом. По истечении определенного времени мастер по наращиванию ресниц проведет небольшую их коррекцию, после чего Вы сможете продолжать радовать своими фантастическими ресницами себя и окружающих. Повторное посещение мастера-специалиста займет значительно меньше времени по сравнению с первым посещением. Реснички можно наращивать, готовясь к особому событию или перед отпуском, но большинство наших клиенток носит их постоянно, периодически корректируя. От такой красоты очень трудно отказаться!
Теперь уже Вам не нужно будет тратить время на ежедневные подкрашивания ресниц тушью и прочими средствами, и беспокоиться об их внешнем виде. По крайней мере, на протяжении месяца и даже более Вы сможете наслаждаться своим бесподобным внешним видом. По истечении определенного времени мастер по наращиванию ресниц проведет небольшую их коррекцию, после чего Вы сможете продолжать радовать своими фантастическими ресницами себя и окружающих. Повторное посещение мастера-специалиста займет значительно меньше времени по сравнению с первым посещением. Реснички можно наращивать, готовясь к особому событию или перед отпуском, но большинство наших клиенток носит их постоянно, периодически корректируя. От такой красоты очень трудно отказаться!
Процедура наращивания представляет собой процесс прикрепления искусственных ресничек с помощью специального клея, изготовленного из натуральных компонентов, на природные ресницы. Можно удлинить каждую ресничку или выборочно в зависимости от желаемой густоты и эффекта.
При наращивании ресниц определенным образом получаются разные эффекты, вот самые распространенные из них.
Натуральный эффект. Классическое наращивание, создающее ресницы натурального объема. Ресницы одной длины и толщины наращиваются от внешнего угла глаза к внутреннему. Реснички подбираются исходя из состояния натуральных ресниц. Наиболее востребованная длина ресниц при таком эффекте — 8, 10 и 11 мм — они позволяют создать естественный эффект длинных от природы ресниц. На самом деле натуральные ресницы длиной 10 мм пусть нечасто, но встречаются. Сюда же можно включить и обычный эффект, при создании которого используются ресницы длиной 6 и 8 мм, то есть самые распространенные природные длины.
Уголки глаз. Пользуется спросом наращивание ресниц в уголках глаз с внешней стороны. В этом случае создается неповторимый эффект как бы отдельно растущих ресниц, когда контрастируют участки с короткими и длинными ресницами. При этом появляется неотразимый и естественный вид, который только усиливается при наличии одинакового оттенка искусственных и натуральных ресниц.
Лисий эффект. Потрясающий эффект создает наращивание под названием лисий взгляд. В этом случае наращиваются реснички, размер которых увеличиваются к внешнему краю глаза. Этот метод делает глаза немного раскосыми и удлиненными с внешней стороны и придает взгляду особую выразительность и загадочность.
Беличий эффект. Ещё один эффект, позволяющий придать взгляду особую чувственность и загадочность- это так называемый беличий эффект. Сам по себе эффект довольно необычен. Это наращивание с удлинением к внешнему углу глаза, похожее на лисий эффект с той разницей, что при процедуре наращивание ресниц беличий эффект создается путем приклеивания ультрадлинных ресниц не на сам уголок глаза, а с отступом к внутренней стороне примерно 5мм. Очень хороший эффект для вечеринки, хотя, в принципе, подходит и для постоянной носки, особенно ультрамодным женщинам.
Наращивание премиум класса (3D эффект) достигается благодаря использованию новых современных материалов премиум класса производства США. Это глянцевые блестящие реснички, они немного толще обычных ресниц и темнее по цвету, что делает взгляд более тёмным и выразительным.
Это глянцевые блестящие реснички, они немного толще обычных ресниц и темнее по цвету, что делает взгляд более тёмным и выразительным.
Мультицветной эффект — это соединение одного или нескольких оттенков, выбор которых полностью зависит от предпочтений потенциальной обладательницы таких ресниц. Цветные реснички будут отлично дополнять наращенные или натуральные ресницы черного цвета. Для такого наращивания также используются стразы, цветной песок и перья. Особой популярностью эффект мультицветного наращивания пользуется в преддверии какого-либо торжественного мероприятия, хотя и в повседневной жизни такие реснички не станут лишними.
Кукольный эффект. Оригинальный и креативный эффект, позволяющий женщине выгодно отличаться от других — это так называемый кукольный эффект. Этот вид наращивания делает глаза огромными, а женщина становится похожа на куколку. Эффект создается с помощью особо длинных ресничек, накладываемых исключительно поресничным способом. Эффект будет незаменим, если девушка собралась на дискотеку, праздничный карнавал, конкурс красоты.
Эффект будет незаменим, если девушка собралась на дискотеку, праздничный карнавал, конкурс красоты.
Разреженный эффект (лучики) — способ наращивания ресниц, последовательно сочетающий две разные длины. При этом создается эффект отдельных длинных ресниц. Также мастера называют его эффектом «лучики». Глаза в этом случае выглядят наиболее естественно, поскольку для такого наращивания используются реснички разной длины. Накладываются они поочередно на определенном расстоянии друг от друга, и выглядят натурально — ведь живые ресницы растут неравномерно и такой эффект в точности повторяет их натуральный рост. Идеально подходит для женщин, которые недовольны густотой и длиной собственных ресниц. Это способ немного изменить свой внешний вид. Такие ресницы подходят для повседневной носки.
- Рекомендации:
- Не окрашивать перед процедурой
- Не наращивать перед менструацией за 5 дней.
- С наращенными ресницами следует обращаться аккуратно: не спать лицом в подушку, не тереть глаза.

- Повышенная чувствительность глаз.
- Повышенное слезоотделение.
- Воспаление век.
Наращивание ресниц «Лисий эффект»
Перед процедурой наращивания ресниц мастер нашего салона красоты внимательно выслушает Ваши пожелания, для того, чтобы иметь возможность предложить наиболее оптимальный для Вас вариант. А также подобрать необходимый Вам цвет ресниц (черный, коричневый, синий и др.).
Способы наращивания
Существует несколько вариантов наращивания:
- Сделать взгляд открытым, т.е. сделать глаза более выразительными. Особенно популярным и часто рекомендуется этот вид наращивания является для тех девушек, у которых есть проблема «нависшего верхнего века», и не только.
- Сделать взгляд «хитрым» поможет наращивание ресниц «лисий эффект». Оно заключается в том, что у внешнего уголка глаз наращиваемые ресницы длиннее, чем средняя часть века и внутренний уголок глаз. Этот вид наращивания очень подходит для девушек с «миндальным» разрезом глаз, т.е. азиаток.
 Этот вид наращивания очень подходит для девушек с «миндальным» разрезом глаз, т.е. азиаток.
Этот вид наращивания очень подходит для девушек с «миндальным» разрезом глаз, т.е. азиаток.Лисий эффект (фото 1)
Лисий эффект (фото 1)
Лисий эффект (фото 2)
Лисий эффект (фото 2)
Лисий эффект (фото 3)
Лисий эффект (фото 3)
Лисий эффект (фото 4)
Лисий эффект (фото 4)
Лисий эффект (фото 5)
Лисий эффект (фото 5)
Лисий эффект (фото 6)
Лисий эффект (фото 6)
Наращивание ресниц заключается в том, что к каждой вашей родной реснице с помощью гипоаллергенной смолы приклеивается искусственная . Как эта искусственная ресничка будет приклеена на вашу ресничку — это уже зависит от квалификации мастера. В нашем салоне это делается на столько профессионально, что даже при близком разглядывании никто не отличит новые ресницы от натуральных!
Противопоказания
| Повышенная чувствительность глаз | Слезоточивость | Блефарит |
Каждая наша ресничка живет 90 дней. В течение этого времени меняется целый ряд ресниц. Ресницы растут в несколько рядов — все разного размера, формы, толщины и изгиб. Когда приходит время выпадать, она выпадает вместе с наращенной . Некоторые паникуют, но зря, это естественный процесс замены ресниц. Наращенные ресницы никакого негативного влияния на наши родные не оказывают. Поэтому 1 раз в 2-4 недели необходимо делать коррекцию. Этот процесс длится около 1,5 часа. Клей со временем разрушается, поэтому сначала снимаются те ресницы, плохо держатся, а потом приклеиваются остальные, где есть необходимость , и у Вас снова роскошный вид.
В течение этого времени меняется целый ряд ресниц. Ресницы растут в несколько рядов — все разного размера, формы, толщины и изгиб. Когда приходит время выпадать, она выпадает вместе с наращенной . Некоторые паникуют, но зря, это естественный процесс замены ресниц. Наращенные ресницы никакого негативного влияния на наши родные не оказывают. Поэтому 1 раз в 2-4 недели необходимо делать коррекцию. Этот процесс длится около 1,5 часа. Клей со временем разрушается, поэтому сначала снимаются те ресницы, плохо держатся, а потом приклеиваются остальные, где есть необходимость , и у Вас снова роскошный вид.
Рекомендуется носить наращенные ресницы 6 месяцев и делать на 1-2 месяца перерыв.
Во время перерыва можно оздоровить и укрепить Ваши реснички благодаря маскам на основе растительных экстрактов и витаминов. Это обеспечит не только достаточное укрепление и питание, но и увеличит их рост, плотность и объем, что поможет вашим ресничкам выглядить как наращенные реснички голливудских звезд.
Лисий эффект наращивания ресниц. Советы по наращиванию на Melady.by
При лисьем эффекте внешний уголок глаза удлиненный и идёт на уменьшение ко внутренней части глаза.
Что делает этот эффект?
Лисий эффект придает хитрость глазам, делает их немного продолговатыми. Идеально этот эффект подойдет девушкам, у которых близко посажены глаза – он визуально их немного «растянет».
Он подойдет для больших и круглых глаз, а также тем, у кого приподняты уголки глаз. Лисий эффект визуально удлиняет глаза и придает гармоничности образу.
Как делать?
Этот эффект желательно выполнять изгибами C и C+. Более сильные изгибы будут смотреться уже менее гармонично. Для выраженного длинного уголка с переходом к коротким ресницам лучше использовать именно изгиб C.
Что сказать по поводу длин?
Мы будем использовать практически все длины, от 6 до 15 мм. При таком наборе длин мы получим наиболее ярко выраженный лисий эффект.
При таком наборе длин мы получим наиболее ярко выраженный лисий эффект.
Если нужен более сдержанный вариант – рекомендуем взять разбег длин поменьше, от 8 до 13 мм. В таком случае лисий эффект также хорошо выражен, но переход к краю глаз более плавный.
Схема лисьего эффекта
Есть два основных вида данного наращивания:
1) Мы равномерно увеличиваем длину от внутреннего уголка глаза ко внешнему и заканчиваем самой большой длиной
2) Длина также увеличивается ко внешнему углу, но самый-самый край глаза идет на резкое уменьшение длины (как на схеме):
Девушкам с маленькими глазами или с опущенными уголками глаз применять данные эффект не рекомендуем, так как в таком случае он визуально слишком сильно опустит внешний угол глаза. Также лучше не делать лисье наращивание девушкам с далеко посаженными глазами.
Примеры фото лисьего эффекта
Подробнее о других эффектах и видах наращивания ресниц читайте здесь
Подобрать нужные реснички для ваших работ можно в каталоге по ссылке
Если у вас остались какие-либо вопросы — обязательно звоните — разберемся вместе! 🙂
Тел. Instagram: @melady.by С уважением, Елена, |
 : +375 33 335-82-10
: +375 33 335-82-10
Интернет-магазин Melady.by — материалы для наращивания ресниц в Беларуси
Сертифицированные
материалы
Регулярные поставки
Доставка по Беларуси
ресничек на клетках, прилегающих к опухолям, могут влиять на передачу сигналов, влияющих на рост рака и ответ на лечение | Онкологический центр Fox Chase
Эрика Големис, PhDPHILADELPHIA (25 мая 2018 г.) — Растущее количество исследований показывает, что реснички, крохотные выросты на поверхности некоторых клеток, играют ключевую роль в определении того, будут ли раковые клетки расти, распространяться и реагировать к терапии. В перспективной статье, посвященной текущим исследованиям, опубликованным в журнале Nature Reviews Cancer , Эрика Големис, доктор философии, заместитель главного научного сотрудника онкологического центра Fox Chase, и соавторы из Китая и России сообщают, что новое понимание ресничек имеет значение. для поведения лекарств от рака и опухолей.
для поведения лекарств от рака и опухолей.
«Как прогрессирование опухоли, так и терапевтический ответ зависят от взаимодействия между раковыми клетками и близлежащими незлокачественными клетками в микроокружении опухоли», — говорится в статье. Големис отметил, что некоторые опухоли и целевые методы лечения рака могут манипулировать клетками, чтобы либо генерировать реснички, либо подавлять их, и что неожиданное отсутствие или присутствие ресничек приводит к запутанной коммуникации между клетками, что поддерживает рост рака.
Согласно перспективе, поскольку реснички выступают во внеклеточное пространство, они позиционируются как пространственно ограниченные центры, которые могут принимать сигналы от других клеток.Некоторые нераковые клетки играют важную роль в канцерогенезе и могут обмениваться информацией с раковыми клетками, чтобы поочередно способствовать росту опухоли, обеспечивать устойчивость к стрессам окружающей среды или лечению рака или поддерживать метастазирование.
Находки на ресничках могут в конечном итоге использоваться для помощи в принятии клинических решений.
«Мы и другие начинаем понимать, что некоторые нацеленные противораковые препараты и химиотерапия изменяют наличие ресничек в клетках в микросреде опухоли», — сказал Големис.«Понимание влияния опухолей и лекарств на реснички потенциально меняет парадигму».
NIH обеспечивает выплату зарплаты доктору Големису.
Включение ресничек: транскрипционные сети, регулирующие цилиогенез | Развитие
Типоспецифические факторы транскрипции клеток могут также действовать через RFX и FOXJ1, чтобы инициировать образование ресничек, при этом независимо регулируя специфические аспекты цилиогенеза для создания разнообразия ресничек (см.рис.5). Например, чтобы создать сотни подвижных ресничек, которые существуют на мультицилийных клетках в дыхательных путях млекопитающих или в эпидермисе Xenopus , сначала необходимо сгенерировать сотни базальных тел, от которых будут отходить аксонемы. Эти базальные тельца возникают de novo из процентриолов, образованных на поверхности дейтеросом — глобулярных структур, которые служат организующими центрами (Anderson, Brenner, 1971; Сорокин, 1968), которые только недавно начали определяться на молекулярном уровне (Klos Dehring et al., 2013; Zhao et al., 2013). Как обсуждалось в предыдущем разделе, FOXJ1 не требуется для образования нескольких базальных телец, но функционирует ниже по течению, при стыковке базальных телец с апикальной клеточной мембраной и последующем удлинении аксонемы (Brody et al., 2000; Gomperts et al. др., 2004; You et al., 2004). Недавно было обнаружено, что белок, содержащий домен спиральной спирали, мультицилин (MCIDAS), необходим для образования мультицилифицированных клеток в эпидермисе Xenopus и дыхательных путях мыши (Stubbs et al., 2012). В обоих контекстах фактор транскрипции MYB действует ниже MCIDAS, генерируя мультицилифицированные клетки. Другой фактор, по-видимому, действует избыточно с MYB, однако, поскольку MYB-дефицитные клетки дыхательных путей обнаруживают задержку, но не полную потерю мульцилиогенеза (Tan et al.
Эти базальные тельца возникают de novo из процентриолов, образованных на поверхности дейтеросом — глобулярных структур, которые служат организующими центрами (Anderson, Brenner, 1971; Сорокин, 1968), которые только недавно начали определяться на молекулярном уровне (Klos Dehring et al., 2013; Zhao et al., 2013). Как обсуждалось в предыдущем разделе, FOXJ1 не требуется для образования нескольких базальных телец, но функционирует ниже по течению, при стыковке базальных телец с апикальной клеточной мембраной и последующем удлинении аксонемы (Brody et al., 2000; Gomperts et al. др., 2004; You et al., 2004). Недавно было обнаружено, что белок, содержащий домен спиральной спирали, мультицилин (MCIDAS), необходим для образования мультицилифицированных клеток в эпидермисе Xenopus и дыхательных путях мыши (Stubbs et al., 2012). В обоих контекстах фактор транскрипции MYB действует ниже MCIDAS, генерируя мультицилифицированные клетки. Другой фактор, по-видимому, действует избыточно с MYB, однако, поскольку MYB-дефицитные клетки дыхательных путей обнаруживают задержку, но не полную потерю мульцилиогенеза (Tan et al. , 2013). MYB также, как было показано, необходим для образования мультицилиндрических клеток в почках рыбок данио (Wang et al., 2013). Основываясь на экспериментах с потерей функции и сверхэкспрессией, MCIDAS и MYB, по-видимому, действуют по единому пути, при этом MCIDAS действует ниже передачи сигналов NOTCH, но выше MYB, чтобы активировать гены, которые управляют формированием нескольких базальных тел [например, Plk4 ( polo-like kinase 4) и Stil (Scl / Tal1 interrupting locus)], с одной стороны, и для включения FOXJ1 для активации генов, необходимых для стыковки базального тела, роста ресничек и подвижности, с другой стороны (Stubbs et al. ., 2012; Tan et al., 2013; Wang et al., 2013). Молекулярные детали этого процесса, такие как прямые мишени MCIDAS и MYB в предшественниках мультицилифицированных клеток, в настоящее время неизвестны. Эта информация будет особенно показательной в случае MCIDAS, который лишен очевидного DBD в своей структуре, но способен локализоваться в ядре и, по-видимому, способен активировать транскрипцию при сверхэкспрессии (Stubbs et al.
, 2013). MYB также, как было показано, необходим для образования мультицилиндрических клеток в почках рыбок данио (Wang et al., 2013). Основываясь на экспериментах с потерей функции и сверхэкспрессией, MCIDAS и MYB, по-видимому, действуют по единому пути, при этом MCIDAS действует ниже передачи сигналов NOTCH, но выше MYB, чтобы активировать гены, которые управляют формированием нескольких базальных тел [например, Plk4 ( polo-like kinase 4) и Stil (Scl / Tal1 interrupting locus)], с одной стороны, и для включения FOXJ1 для активации генов, необходимых для стыковки базального тела, роста ресничек и подвижности, с другой стороны (Stubbs et al. ., 2012; Tan et al., 2013; Wang et al., 2013). Молекулярные детали этого процесса, такие как прямые мишени MCIDAS и MYB в предшественниках мультицилифицированных клеток, в настоящее время неизвестны. Эта информация будет особенно показательной в случае MCIDAS, который лишен очевидного DBD в своей структуре, но способен локализоваться в ядре и, по-видимому, способен активировать транскрипцию при сверхэкспрессии (Stubbs et al. , 2012).
, 2012).
Формирование подвижных моноцилий в вентральном узле эмбриона мыши программируется с помощью фактора транскрипции гомеобокса, называемого НОТО (гомеобокс хорды).Мыши, лишенные NOTO, обнаруживают укороченные и деформированные узловые реснички и последующее нарушение лево-правой асимметрии (Beckers et al., 2007). Чтобы сгенерировать моноцилии узла, NOTO развертывает FOXJ1, который, в свою очередь, активирует Rfx3 . В элегантном эксперименте Альтен и его коллеги заменили кодирующую область мыши Noto на Foxj1 ( Noto :: Foxj1 ) и обнаружили, что, в отличие от мутантов Noto , экспрессия генов ресничек (включая Rfx3 ) был восстановлен, а длина и подвижность ресничек были восстановлены до нормальных значений (Alten et al., 2012). Однако эти эмбрионы продолжали демонстрировать дефекты лево-правой асимметрии, поскольку поляризованная ориентация узловых ресничек, которая зависит от пути планарной полярности клеток (PCP) (Hashimoto et al. , 2010; Song et al., 2010), оставалась нарушенной. у мышей Noto :: Foxj1 . Таким образом, NOTO, по-видимому, активирует Foxj1 и Rfx3 для генерации аксонем подвижных узловых ресничек, независимо устанавливая их правильное заднее положение на узловых клетках, возможно, путем активации пути PCP (Alten et al., 2012).
, 2010; Song et al., 2010), оставалась нарушенной. у мышей Noto :: Foxj1 . Таким образом, NOTO, по-видимому, активирует Foxj1 и Rfx3 для генерации аксонем подвижных узловых ресничек, независимо устанавливая их правильное заднее положение на узловых клетках, возможно, путем активации пути PCP (Alten et al., 2012).
В Drosophila пронейральный транскрипционный фактор Atonal (ATO) управляет дифференцировкой хордотональных нейронов. Как часть этой программы, ATO активирует экспрессию как fd3F , так и Rfx , чтобы генерировать подвижные механосенсорные реснички, которые отходят от этих нейронов (Cachero et al., 2011). Интересно, что ATO, как было показано, непосредственно активирует экспрессию цилиарного компонента, Dilatory / CEP131, указывая на то, что ATO может вносить вклад в цилиогенез независимо от кассеты RFX / FD3F (Cachero et al., 2011; Ма и Джарман, 2011). Действительно, эта функция ATO может представлять собой фундаментальный механизм образования механосенсорных ресничек, потому что у рыбок данио ортолог ATO (ATOh2B) активирует паралог foxj1 в волосковых клетках внутреннего уха, что приводит к образованию неподвижных киноцилий ( Ю. и др., 2011).
Действительно, эта функция ATO может представлять собой фундаментальный механизм образования механосенсорных ресничек, потому что у рыбок данио ортолог ATO (ATOh2B) активирует паралог foxj1 в волосковых клетках внутреннего уха, что приводит к образованию неподвижных киноцилий ( Ю. и др., 2011).
Редокс-регуляция подвижных ресничек при заболеваниях дыхательных путей
https://doi.org/10.1016/j.redox.2019.101146Получение прав и содержаниеРезюме
Подвижные реснички на клетках дыхательных путей необходимы для выведения захваченных слизью частиц из легкое.Ресничные эпителиальные клетки дыхательных путей подвергаются уникальному воздействию окислителей за счет улавливания частиц, мусора и патогенов в слизи и прямого воздействия вдыхаемых газов-окислителей. Динеин-АТФазы, двигатели, управляющие подвижностью ресничек, чувствительны к локальной окислительно-восстановительной среде внутри каждой реснички. Несколько белков, локализованных в ресничках, чувствительных к окислению-восстановлению, модулируют активность динеина и включают протеинкиназу A, протеинкиназу C и протеинфосфатазу 1. Более того, реснички богаты известными окислительно-восстановительными регуляторными белками и белками, содержащими тиоредоксиновый домен, которые имеют решающее значение для поддержания сбалансированного окислительно-восстановительного потенциала. окружающая обстановка.Важно отметить, что нонсенс-мутация в TXNDC3, которая содержит мотив тиоредоксина, была недавно идентифицирована как вызывающая заболевание при первичной цилиарной дискинезии, наследственном заболевании подвижных ресничек, приводящем к нарушению мукоцилиарного клиренса. Здесь мы делаем обзор современного понимания роли (й) окислителей в изменении функции ресничек дыхательных путей. Мы фокусируемся на окислителях, образующихся в дыхательных путях, окислительно-восстановительных целях ресничек, которые модулируют биение ресничек, и дисбалансе окислительно-восстановительного состояния, влияющем на здоровье и болезни.Наконец, мы рассматриваем модели болезней, такие как курение, астма, употребление алкоголя и инфекции, а также прямое применение оксидантов, которые влияют на окислительно-восстановительный баланс как модулятор подвижности ресничек.
Более того, реснички богаты известными окислительно-восстановительными регуляторными белками и белками, содержащими тиоредоксиновый домен, которые имеют решающее значение для поддержания сбалансированного окислительно-восстановительного потенциала. окружающая обстановка.Важно отметить, что нонсенс-мутация в TXNDC3, которая содержит мотив тиоредоксина, была недавно идентифицирована как вызывающая заболевание при первичной цилиарной дискинезии, наследственном заболевании подвижных ресничек, приводящем к нарушению мукоцилиарного клиренса. Здесь мы делаем обзор современного понимания роли (й) окислителей в изменении функции ресничек дыхательных путей. Мы фокусируемся на окислителях, образующихся в дыхательных путях, окислительно-восстановительных целях ресничек, которые модулируют биение ресничек, и дисбалансе окислительно-восстановительного состояния, влияющем на здоровье и болезни.Наконец, мы рассматриваем модели болезней, такие как курение, астма, употребление алкоголя и инфекции, а также прямое применение оксидантов, которые влияют на окислительно-восстановительный баланс как модулятор подвижности ресничек.
Ключевые слова
S -нитрозирование
Спирт
Реснички
Редокс-регулирование
Пероксид водорода
Супероксид
Оксид азота
Сокращения
AICD, вызванный алкоголем, CBO,
частота биений ресничек
DTNB5,5′-дитио-бис- [2-нитробензойная кислота]
FiO 2фракция вдыхаемого кислорода
Hsp90белок теплового шока 90
l-NAMEметиловый эфир N-нитроаргинина
NOXникатиномид-аденин-динуклеотидфосфат-оксидаза
Nrf2Ядерный фактор (эритроидный 2) -подобный 2
PCDпервичная цилиарная дискинезия
PP2Aпротеинфосфатаза 2A
RHAMM рецепторRHAMM
, связанный с кислородом, рецептор
RHAMM
, связанный с гиалуроновой кислотой моалуроновой кислоты
для гиалуроновой кислоты
виды
RSVреспираторно-синцитиальный вирус
sGCрастворимая гуанилилциклаза
TrxR2 900 02 тиоредоксинредуктаза 2 TXNDC, содержащий тиоредоксиновый домен
XDH / XOксантиндегидрогеназа / ксантиноксидаза
Рекомендуемые статьиЦитирующие статьи (0)
© 2019 Авторы. Опубликовано Elsevier B.V.
Рекомендуемые статьи
Цитирующие статьи
Путь киназы LRRK2 болезни Паркинсона для блокирования первичных ресничек и передачи сигналов Sonic hedgehog в головном мозге
[ От редакции: Эта статья прошла редакционный процесс, в ходе которого авторы решают, как реагировать на вопросы, поднятые во время рецензирования. По оценке редактора-рецензента, все проблемы решены.]
Благодарим вас за отправку вашей статьи «Путь киназы LRRK2 болезни Паркинсона для блокирования первичных ресничек и передачи сигналов Sonic hedgehog в головном мозге» на рассмотрение eLife .Ваша статья была рассмотрена тремя рецензентами, включая Кристофера Берда в качестве редактора-рецензента и рецензента №1, а оценку контролировал Вивек Малхотра в качестве главного редактора. Следующее лицо, участвовавшее в рассмотрении вашей заявки, согласилось раскрыть свою личность: Питер Новик (Рецензент № 3). Рецензент №2 остается анонимным.
Редактор-рецензент выделил проблемы, требующие доработки и / или ответов, и мы включили отдельные обзоры ниже для вашего рассмотрения.Если у вас есть какие-либо вопросы, не стесняйтесь обращаться к нам.
Резюме:
Рукопись представляет исследование ролей LRRK2, Rab8A, Rab10 и RILPL1 в цилиогенезе и сообщает, что связанные с болезнью Паркинсона изменения активности LRRK2 приводят к нарушению сборки ресничек и передаче сигналов Hedgehog. В целом, авторы обзора сочли вывод, что LRRK2 регулирует активность Rab8 и Rab10 в цилиогенезе, хорошо подтверждается данными. Хотя исследование дает мало механистического понимания специфических функций Rab8 и Rab10 в цилиогенезе, оно действительно дает интересное и потенциально важное понимание этиологии болезни Паркинсона.
Основные проблемы:
1) Точно не установлено, что Rab8 и Rab10 играют важную роль в цилиогенезе в типах клеток, отличных от клеточной линии A549 (см. Обзор 2, главный комментарий 1), который обычно не используется для исследования цилиогенеза. Влияние работы можно было бы лучше оценить, повторив ограниченное количество ключевых экспериментов на клетках RPE или MEF дикого типа (например, чтобы показать, что Rab10 / RILPL1 может ингибировать цилиогенез в этих клетках). По-видимому, это было решено с помощью экспериментов, проведенных с использованием клеток RPE (подраздел «Rab8 активирует, в то время как Rab10 ингибирует образование ресничек в клетках A549», первый параграф), но данные не показаны.Как минимум, эти данные должны быть показаны.
Влияние работы можно было бы лучше оценить, повторив ограниченное количество ключевых экспериментов на клетках RPE или MEF дикого типа (например, чтобы показать, что Rab10 / RILPL1 может ингибировать цилиогенез в этих клетках). По-видимому, это было решено с помощью экспериментов, проведенных с использованием клеток RPE (подраздел «Rab8 активирует, в то время как Rab10 ингибирует образование ресничек в клетках A549», первый параграф), но данные не показаны.Как минимум, эти данные должны быть показаны.
2) Ни Rab8, ни Rab10 мутантные белки T-to-E / A не являются функциональными, и кажется вероятным, что мутантные Rab нефункциональны, независимо от их предполагаемого воздействия на фосфорилирование, и этот момент недостаточно изучен или четко не передан. Показано, что Rab10 T73A является цитозольным, что объясняет его нефункциональность. Почему другие мутанты rab8 и Rab10 нефункциональны? Они изопренилированы? Если мутанты не влияют избирательно на фосфорилирование (например,г. они изменяют изопренилирование), их использование кажется несущественным по отношению к основной теме статьи.
3) Следует определить влияние мутаций сайта фосфорилирования Rab10 LRRK2 (T73A, E) на его ассоциацию с RILP1, аналогично анализу мутантов Rab8 и RILP1 (Рисунок 3 — рисунок в приложении 1). Эта информация важна для интерпретации фенотипов мутантных белков.
4) Обратите внимание, что каждого из рецензентов смущали определенные аспекты презентации (см. Обзоры).Я призываю вас рассмотреть возможность пересмотра презентации, чтобы устранить эти источники путаницы.
Отдельных отзывов (ответьте по каждому пункту):
Рецензент № 1:
В этом исследовании рассматривается роль LRRK, протеинкиназы, участвующей в болезни Паркинсона, в цилиогенезе. Мутации в LRRK, вызывающие БП, активируют его протеинкиназную активность, но как это приводит к БП, неизвестно. Исследование следует за более ранней публикацией, в которой сообщалось, что белки Rab являются основными субстратами для LRRK, включая Rab8 и Rab10, которые являются предметом настоящего исследования. Rab8 ранее участвовал в цилиогенезе, и авт. Представляют доказательства, что фосфорилирование Rab8 с помощью LRRK ингибирует его функцию в цилиогенезе. Напротив, авт. Обнаруживают, что истощение Rab10, другой Rab GTPase, участвующей в цилиогенезе, потенцирует цилиогенез и что фосфорилирование Rab10 с помощью LRRK усиливает его ингибирующее действие на цилиогенез посредством усиленного взаимодействия с RILPL1, эффектором Rab10 с неизвестной функцией. Показано, что истощение RILPL1 потенцирует цилиогенез, подобно истощению Rab10.Наконец, показано, что мутации PD в LRRK вызывают снижение доли реснитчатых клеток и недостаточную передачу сигналов SSH в холинергических нейронах полосатого тела мозга мышей. Специфические функции Rab8 или Rab10 в цилиогенезе или PD в значительной степени неизвестны. Это технически надежное, хорошо контролируемое исследование, которое четко подтверждает роли Rab8 и Rab10 в цилиогенезе, хотя, если эти GTPases играют прямую роль, конкретные роли в цилиогенезе остаются открытым вопросом.
Rab8 ранее участвовал в цилиогенезе, и авт. Представляют доказательства, что фосфорилирование Rab8 с помощью LRRK ингибирует его функцию в цилиогенезе. Напротив, авт. Обнаруживают, что истощение Rab10, другой Rab GTPase, участвующей в цилиогенезе, потенцирует цилиогенез и что фосфорилирование Rab10 с помощью LRRK усиливает его ингибирующее действие на цилиогенез посредством усиленного взаимодействия с RILPL1, эффектором Rab10 с неизвестной функцией. Показано, что истощение RILPL1 потенцирует цилиогенез, подобно истощению Rab10.Наконец, показано, что мутации PD в LRRK вызывают снижение доли реснитчатых клеток и недостаточную передачу сигналов SSH в холинергических нейронах полосатого тела мозга мышей. Специфические функции Rab8 или Rab10 в цилиогенезе или PD в значительной степени неизвестны. Это технически надежное, хорошо контролируемое исследование, которое четко подтверждает роли Rab8 и Rab10 в цилиогенезе, хотя, если эти GTPases играют прямую роль, конкретные роли в цилиогенезе остаются открытым вопросом. На мой взгляд, основная ценность этого исследования заключается в идентификации функциональных последствий для LRRK-обеспечиваемого фосфорилирования Rab8 и Rab10 в цилиогенезе, открытие, которое проливает свет на цилиогенез для будущих исследований этиологии PD.
На мой взгляд, основная ценность этого исследования заключается в идентификации функциональных последствий для LRRK-обеспечиваемого фосфорилирования Rab8 и Rab10 в цилиогенезе, открытие, которое проливает свет на цилиогенез для будущих исследований этиологии PD.
Фенотипический анализ мышей с нокаутом Rab8a / b, проведенный Sato et al., 2014, показал по существу не влияние на количество ресничек и мало свидетельств цилиопатий, которых можно было бы ожидать из-за потери ключевого фактора цилиогенеза. Более того, они сообщили, что нокаут Rab10 приводит к потере ресничек, а здесь сообщается об обратном. Как авторы согласовывают эти опубликованные наблюдения со своими результатами? Если я правильно понимаю, авт. Предполагают, что Rab8 «активирует» цилиогенез, а не выполняет существенную роль, но мне неясно, что означает «активировать».
Показано, что потеря RILPL1 усиливает цилиогенез, подобно потере Rab10. Также показано, что Rab10 не требует RILPL1 для его эффектов на цилиогенез («Это предполагает, что Rab8A и Rab10 не нуждаются в RILPL1 для их эффектов на цилиогенез…»). Так почему же предполагается, что Rab10 и RILPL1 («киназа LRRK2 действует через путь pRab10-RILPL1, чтобы ингибировать цилиогенез») функционировали по одному и тому же пути цилиогенеза? Было бы полезно, если бы авторы прояснили этот вывод.
Так почему же предполагается, что Rab10 и RILPL1 («киназа LRRK2 действует через путь pRab10-RILPL1, чтобы ингибировать цилиогенез») функционировали по одному и тому же пути цилиогенеза? Было бы полезно, если бы авторы прояснили этот вывод.
Файлы дополнительных данных и статистические комментарии:
Достоверность и объем статистической информации удовлетворительны.
Рецензент № 2:
Здесь Dhekne et al. исследовать роль LRRK2, Rab8A, Rab10 и RILPL1 в цилиогенезе и сообщить, что связанные с болезнью Паркинсона изменения активности LRRK2 приводят к нарушению сборки ресничек и передачи сигналов Hedgehog, что может вносить вклад в патогенез Паркинсона. Это исследование основано на двух недавних отчетах в eLife из этих групп (работающих также с группой Матиаса Манна), которые идентифицировали Rab8A и Rab10 как субстраты LRRK2, охарактеризовали зависимое от фосфорилирования связывание RILPL1 с этими Rab и обнаружили снижение цилиогенеза в фибробластах с повышенным уровнем цилиогенеза. LRRK2 активность.В этой рукописи авт. Исследуют, как Rab8A, Rab10 и RILPL1 контролируют цилиогенез ниже LRRK2, обнаружив, что Rab8A способствует цилиогенезу (как сообщалось ранее), тогда как RILPL1 способствует Rab10-зависимому ингибированию образования ресничек. Кроме того, авт. Показывают, что повышенная активность LRRK2 ведет к снижению сниженного цилиогенеза (в MEFs, человеческих iPS клетках и мозге мышей) и к снижению зависимой от ресничек передачи сигналов Hedgehog (в MEFs и человеческих iPS клетках).
LRRK2 активность.В этой рукописи авт. Исследуют, как Rab8A, Rab10 и RILPL1 контролируют цилиогенез ниже LRRK2, обнаружив, что Rab8A способствует цилиогенезу (как сообщалось ранее), тогда как RILPL1 способствует Rab10-зависимому ингибированию образования ресничек. Кроме того, авт. Показывают, что повышенная активность LRRK2 ведет к снижению сниженного цилиогенеза (в MEFs, человеческих iPS клетках и мозге мышей) и к снижению зависимой от ресничек передачи сигналов Hedgehog (в MEFs и человеческих iPS клетках).
Негативная регуляция цилиогенеза с помощью Rab10 / RILPL1 — интересная находка, которая, если ее можно обобщить на др. Типы клеток / ткани, представляет новое понимание цилиогенеза, которое мотивирует будущий анализ лежащего в основе механизма.Кроме того, находки, что гиперактивность LRRK2 ингибирует цилиогенез и притупляет передачу сигналов Hedgehog, предоставляют дополнительные доказательства того, что дефекты ресничек могут вносить вклад в болезнь Паркинсона. Наконец, наблюдение, что нокдаун Rab10 подавляет дефекты цилиогенеза, наблюдаемые в MEFs с повышенной активностью LRRK2, обеспечивает важную связь между двумя вышеупомянутыми открытиями. Как подробно описано ниже, исследование может быть усилено дальнейшим изучением роли Rab10 в цилиогенезе в клеточных линиях, которые чаще используются для изучения ресничек.Кроме того, некоторые данные представлены в запутанной форме и не подтверждают некоторые сделанные выводы. Эти неясные разделы значительно затрудняют понимание основных результатов и должны быть отредактированы перед публикацией в eLife .
Как подробно описано ниже, исследование может быть усилено дальнейшим изучением роли Rab10 в цилиогенезе в клеточных линиях, которые чаще используются для изучения ресничек.Кроме того, некоторые данные представлены в запутанной форме и не подтверждают некоторые сделанные выводы. Эти неясные разделы значительно затрудняют понимание основных результатов и должны быть отредактированы перед публикацией в eLife .
1) Многие эксперименты проводятся на клетках A549, но клетки A549 инфузируют с относительно низкой скоростью (~ 25%) и обычно не используются для изучения цилиогенеза. Таким образом, возможно, что ингибирующие эффекты Rab10 / RILPL1 на цилиогенез могут быть отличительной чертой этого типа клеток, а не свидетельствовать о более общей роли этих белков в регуляции образования ресничек (особенно как Sato et al., 2014, сообщили, что Rab10 вместо этого является позитивным регулятором цилиогенеза, который действует дублирующе с Rab8A и Rab8B). Эта проблема частично смягчается данными по MEF LRRK2-R1441G, показанными на рисунке 6, но, тем не менее, было бы полезно повторить ограниченное количество ключевых экспериментов с клетками RPE или MEF дикого типа (например, чтобы показать, что Rab10 / RILPL1 может ингибировать цилиогенез в эти клетки). В первом абзаце подраздела «Rab8 активирует, в то время как Rab10 ингибирует образование ресничек в клетках A549», авторы отмечают, что реснички были исследованы в клетках RPE после нокдауна Rab8A и Rab10, но данные не показаны.
В первом абзаце подраздела «Rab8 активирует, в то время как Rab10 ингибирует образование ресничек в клетках A549», авторы отмечают, что реснички были исследованы в клетках RPE после нокдауна Rab8A и Rab10, но данные не показаны.
2) Некоторые разделы результатов сбивают с толку и должны быть исправлены (подробности см. В дополнительных комментариях 1-5).
3) Хотя авторы протестировали функциональную взаимозависимость между LRRK2, Rab8A, Rab10 и RILPL1, общая картина того, как эти белки взаимодействуют для контроля цилиогенеза, остается неясной в отношении того, где подходит Rab8A. Могут ли авторы интегрировать Rab8A в путь модель, как на рисунке 6D? Кажется, что одна из возможных моделей — это RILPL1 — | Rab8A — | Rab10 — | ciliogenesis, при этом LRRK2 способствует активности Rab10, но это немного отличается от рис. 6D и, возможно, есть альтернативы, которые подходят лучше или иначе заслуживают рассмотрения.Наконец, учитывая доказательства того, что Rab10 может ингибировать цилиогенез в отсутствие RILPL1, существуют ли эффекторы Rab10, которые авторы рассматривают как хороших кандидатов для обеспечения ингибирования Rab10 цилиогенеза?
Незначительные комментарии:
1) Авторы заявляют, что «добавление экзогенного Rab10 подавляет образование ресничек в клетках, лишенных Rab8A или Rab10. Более того, Rab8A усиливает образование ресничек в клетках, лишенных Rab10; таким образом, это представляет собой независимый путь активации цилиогенеза». Во-первых, влияние экзогенной экспрессии Rab10 на цилиогенез в Rab8A-дефицитных клетках, по-видимому, здесь не исследовалось.Во-вторых, данные показывают, что Rab8A не усиливает образование ресничек в клетках, лишенных Rab10; скорее, фиг. 2B показывает, что клетки, лишенные Rab10, обходят нормальную роль Rab8A в стимулировании цилиогенеза — само по себе поразительное открытие. Наконец, этот раздел завершается обсуждением данных Sato et al. что у мышей с двойным мутантом Rab8A / B обнаружены нормальные первичные реснички. Этот факт, кажется, сбивает с толку или противоречит имеющимся данным; Вместо этого может быть полезно обсудить выводы Sato et al., 2014, в разделе «Обсуждение».
Более того, Rab8A усиливает образование ресничек в клетках, лишенных Rab10; таким образом, это представляет собой независимый путь активации цилиогенеза». Во-первых, влияние экзогенной экспрессии Rab10 на цилиогенез в Rab8A-дефицитных клетках, по-видимому, здесь не исследовалось.Во-вторых, данные показывают, что Rab8A не усиливает образование ресничек в клетках, лишенных Rab10; скорее, фиг. 2B показывает, что клетки, лишенные Rab10, обходят нормальную роль Rab8A в стимулировании цилиогенеза — само по себе поразительное открытие. Наконец, этот раздел завершается обсуждением данных Sato et al. что у мышей с двойным мутантом Rab8A / B обнаружены нормальные первичные реснички. Этот факт, кажется, сбивает с толку или противоречит имеющимся данным; Вместо этого может быть полезно обсудить выводы Sato et al., 2014, в разделе «Обсуждение».
2) Выводы, которые можно сделать на основании использования мутантов T-> A и T-> E по фосфо-сайту, также не ясны. Как отмечают авторы, Rab10A T73A, по-видимому, нефункционален и не должен использоваться в качестве аллеля, блокирующего фосфорилирование. Учитывая, что оба мутанта Rab8A T72A и T72E не способны восстанавливать дефекты цилиогенеза в нокаутных Rab8A клетках и плохо взаимодействуют с RILPL1 (в то время как фосфорилированный Rab8A надежно связывает RILPL1), вероятно, эти мутанты нефункциональны и не специфически блокируют или имитируют фосфорилирование.Таким образом, утверждение в Abstract, что «фосфорилирование Rab8A блокирует его способность способствовать цилиогенезу», должно быть пересмотрено. Кроме того, является ли более сильное ингибирование цилиогенеза мутантом Rab10 T73E по сравнению с Rab10 дикого типа статистически значимым?
Учитывая, что оба мутанта Rab8A T72A и T72E не способны восстанавливать дефекты цилиогенеза в нокаутных Rab8A клетках и плохо взаимодействуют с RILPL1 (в то время как фосфорилированный Rab8A надежно связывает RILPL1), вероятно, эти мутанты нефункциональны и не специфически блокируют или имитируют фосфорилирование.Таким образом, утверждение в Abstract, что «фосфорилирование Rab8A блокирует его способность способствовать цилиогенезу», должно быть пересмотрено. Кроме того, является ли более сильное ингибирование цилиогенеза мутантом Rab10 T73E по сравнению с Rab10 дикого типа статистически значимым?
3) Порядок данных, представленных в тексте, не совсем соответствует порядку данных на рисунках, особенно для первых двух рисунков и рисунка 3 — дополнения к рисунку 1. Регулировка порядка панелей рисунков упростит работу с бумагой. следить.Кроме того, в первом абзаце подраздела «RILPL1 регулирует локализацию pRab10» ссылка на рисунок 3 — рисунок — дополнение 1 вместо ссылки на рисунок 5 — рисунок — приложение 1. В первом абзаце подраздела «RILPL1 регулирует pRab10» локализация », ссылка на рисунок 4B, по-видимому, вместо ссылки на рисунок 4A. Наконец, что показано на рисунке 2A в отношении GFP-Rab8 и GFP-Rab10?
В первом абзаце подраздела «RILPL1 регулирует pRab10» локализация », ссылка на рисунок 4B, по-видимому, вместо ссылки на рисунок 4A. Наконец, что показано на рисунке 2A в отношении GFP-Rab8 и GFP-Rab10?
4) Дополнительная область путаницы в разделе, озаглавленном «RILPL1 подавляет образование ресничек посредством центриолей».Из данных на Рис. 5F-H кажется, что вопреки названию раздела, RILPL1 подавляет образование ресничек независимо от эффектов на расстояние между центриолями. Кроме того, хотя эффекты на цилиогенез экспрессии RILPL1 в RILPL1 KO клетках поразительны, было бы информативно оценить, происходит ли подобное ингибирование цилиогенеза, когда RILPL1 сверхэкспрессируется в клетках дикого типа.
5) На рисунке 4 авторы анализируют влияние фрагментов RILPL1 на локализацию pRab10. Хотя сообщенные эффекты интересны, предположение о том, что N-конец RILPL1 взаимодействует с pRab10 в перицентриолярных мембранах, возможно, следует смягчить, учитывая, что предыдущий отчет этих групп показал, что именно C-концевой RH домен в RILPL1 опосредует взаимодействие с Rab8A (PMID: 2
62), и то же самое может быть верно для Rab10 (даже с учетом показанных данных).
6) Результаты для iPS-клеток LRRK2-G2019S на фиг. 7C-E можно отнести к различиям в цилиогенезе, связанном с процессом получения мутантных линий по сравнению с линиями дикого типа. Это потенциальное беспокойство можно уменьшить, исследуя, восстанавливает ли обработка MLi2 нормальные уровни образования ресничек в клетках G2019S.
7) Авт. Утверждают, что «Yoshimura et al., 2007 были первыми, кто показал, что Rab8A, но не Rab8B, является единственной Rab GTPase, которая локализуется в первичных ресничках в клетках hTert-RPE1».Это утверждение может ввести в заблуждение некоторых читателей, учитывая последующие доказательства того, что GFP-Rab23 локализуется в ресничках RPE1 (PMID: 26136363), как и Rab8B (PMID: 21273506). Кроме того, хотя Rab8 и Rabin8 способствуют цилиогенезу, этот эффект, по-видимому, только частично осуществляется через BBSome и вместо этого может быть в первую очередь связан с доставкой мембранных везикул к растущей ресничке (например, PMID: 25812525).
Рецензент № 3:
В этой статье оценивается роль Rab8 и Rab10 GTPases, а также Rab эффекторного RILPL1 в регуляции цилиогенеза с помощью киназы LRRK2. Затем он переходит к изучению роли изменений в цилиогенезе в опосредовании эффектов LRRK2 при болезни Паркинсона. В целом исследования хорошо спланированы и хорошо выполнены, а результаты в целом убедительны. Хотя статья, вероятно, открывает столько же вопросов, сколько и закрывает, она представляет собой важный шаг вперед в понимании одного молекулярного механизма, лежащего в основе генетической причины болезни Паркинсона, и должна иметь широкую аудиторию. Ниже я выделил ряд моментов, которые следует изучить более тщательно или, в одном случае, исключить для ясности.
Затем он переходит к изучению роли изменений в цилиогенезе в опосредовании эффектов LRRK2 при болезни Паркинсона. В целом исследования хорошо спланированы и хорошо выполнены, а результаты в целом убедительны. Хотя статья, вероятно, открывает столько же вопросов, сколько и закрывает, она представляет собой важный шаг вперед в понимании одного молекулярного механизма, лежащего в основе генетической причины болезни Паркинсона, и должна иметь широкую аудиторию. Ниже я выделил ряд моментов, которые следует изучить более тщательно или, в одном случае, исключить для ясности.
1) Каков уровень экспрессии Rab8 и 10 и их мутантных аллелей на рис. 1D и E по отношению к эндогенному уровню? Смотрится очень высоко.
2) Отношения Rab8 и Rab10 глубоко не исследовались. Влияет ли потеря или избыточная экспрессия одного на локализацию другого в ресничках? Влияет ли потеря или избыточная экспрессия одного на активацию другого? Они тесно связаны и оба могут связывать RILPL, как каждый влияет на взаимодействие другого с этим общим эффектором? Они интерпретируют свои данные, чтобы указать, что эффекты каждого из них на цилиогенез полностью независимы от другого, но я в этом не уверен.
3) На рис. 2B, C и D требуется контроль WT.
4) В тексте ячейки, в которых CRISPR выбивает компонент, называются «истощенными». Точнее было бы сказать, что компонент удален. Истощение предполагает частичную утрату.
5) На рисунке 2E они также должны проверить мембранную ассоциацию мутантов Rab8. Непосредственное тестирование на изопренилирование различных аллелей Rab8 и 10 могло бы решить эту проблему более четко.
6) Рисунок 3 — рисунок в приложении 1 рассматривает влияние фосфорилирования на взаимодействие Rab8 с RILPL, однако эффекты на цилиогенез преимущественно опосредуются Rab10.Им также следует изучить эффекты фосфорилирования Rab10 на его взаимодействие с RILPL.
7) Обсуждение влияния RILPL на центриолярное расстояние очень сбивает с толку и не имеет очевидного отношения к этой истории. На рис. 5G и H похоже, что эффект есть, но на рис. 5I значительного эффекта нет. Это смущает. Я бы предложил исключить этот раздел статьи, поскольку он отвлекает от основной линии рассуждений.
статей PLOS рассматривают заболевание ресничек у собак, бешенство, связанное с лисами, еще
В PLOS Genetics исследователи из Швейцарии, США, Финляндии и Великобритании описывают делецию единственного основания в гене NME5, которая, по-видимому, способствует первичной цилиарная дискинезия (PCD) у аляскинских маламутов. Команда использовала комбинацию полногеномного секвенирования, сцепления и картирования гомозиготности у полдюжины аляскинских маламутов из семьи, пораженной PCD, редким наследственным генетическим заболеванием, которое влияет на подвижные реснички, что приводит к хроническим инфекциям дыхательных путей, проблемам с фертильностью, и другие симптомы.Основываясь на результатах у двух щенков с PCD и их здоровых родителей и братьев и сестер, по сравнению с доступными последовательностями от более чем 600 здоровых собак, авторы сузились до аутосомно-рецессивно наследуемой делеции NME5, которая впоследствии была протестирована на дополнительных собаках с или без PCD и мышиная модель болезни. «Наши результаты позволяют проводить генетическое тестирование на собаках и идентифицировать NME5 как новый ген-кандидат для нераскрытых случаев PCD у человека», — пишут они.
«Наши результаты позволяют проводить генетическое тестирование на собаках и идентифицировать NME5 как новый ген-кандидат для нераскрытых случаев PCD у человека», — пишут они.
Команда Канадского агентства по надзору за пищевыми продуктами отслеживает перемещение вирусов бешенства, связанных с песцами, в провинции Онтарио для статьи в PLOS Neglected Tropical Diseases .Пытаясь понять возобновление случаев бешенства у скунсов на юго-западе Онтарио в 2015 году после трехлетнего перерыва, исследователи проанализировали новые и доступные полногеномные последовательности для 128 изолятов вируса бешенства, собранных в Онтарио в период с 1990-х годов по настоящее время. Их филогенетический анализ выступил против недавнего нового введения вирусов бешенства линии, связанной с песцами. Вместо этого авторы заключают, что «вариант, ответственный за текущие случаи в юго-западном Онтарио, развился из тех вариантов, которые, как известно, циркулировали в Онтарио ранее», предполагая, что «устойчивость бешенства в дикой природе оставалась незамеченной в исследовании в течение почти трех лет. «
«
В статье в PLOS One исследователи из Японии сообщают о потенциальных маркерах микроРНК для лекарственного ответа у пациентов с метастатическим раком молочной железы, получавших химиотерапевтический препарат эрибулин (продаваемый Eisai как Halaven) после предшествующей терапии антрациклином и таксаном. Использование miRNA. Используя массивы и моделирование прогнозов, команда провела поиск информативных miRNA в образцах сыворотки крови, собранных до и после лечения эрибулином у 147 пациентов с метастатическим раком молочной железы, сравнивая профили miRNA у 95 пациентов, у которых в среднем не было новых метастазов в течение 117 месяцев наблюдения. , с 52 пациентами, которые этого не сделали.Восемь сывороточных miRNAs имели очевидную связь с новыми отдаленными метастазами, сообщают авторы, а две из них — miR-8089 и miR-5698 — оказались значительно связаны с общей выживаемостью у пациентов с метастатическим раком груди, получавших эрибулин.
Структурная основа того, как действуют подвижные реснички | Бионаука
Абстрактные
Подвижная ресничка — это чудо механики, клеточная наномашина, которая производит высокоскоростные биения на основе цикла изгибов, которые движутся вдоль аксонемы, состоящей из 9 + 2 микротрубочек. Молекулярные двигатели, динеины, приводят в действие биение ресничек. Динеины компактируются во внутреннюю и внешнюю ветви динеина, активность которых строго регулируется, вызывая скольжение микротрубочек и изгиб аксонемы. Гипотеза точки переключения была разработана давно, чтобы объяснить, как скольжение в присутствии взаимодействий аксонемной радиальной спицы и центральной пары вызывает биение ресничек. С тех пор новая генетическая, биохимическая и структурная сложность была обнаружена, в частности, с мутантами Chlamydomonas, с помощью высокоскоростного анализа движения с высоким разрешением и с помощью криоэлектронной томографии.Мы стоим на пороге новых открытий, касающихся молекулярного контроля подвижности, которые расширяют и уточняют наше понимание основных событий, лежащих в основе переключения активности рук, а также формирования и распространения изгибов.
Молекулярные двигатели, динеины, приводят в действие биение ресничек. Динеины компактируются во внутреннюю и внешнюю ветви динеина, активность которых строго регулируется, вызывая скольжение микротрубочек и изгиб аксонемы. Гипотеза точки переключения была разработана давно, чтобы объяснить, как скольжение в присутствии взаимодействий аксонемной радиальной спицы и центральной пары вызывает биение ресничек. С тех пор новая генетическая, биохимическая и структурная сложность была обнаружена, в частности, с мутантами Chlamydomonas, с помощью высокоскоростного анализа движения с высоким разрешением и с помощью криоэлектронной томографии.Мы стоим на пороге новых открытий, касающихся молекулярного контроля подвижности, которые расширяют и уточняют наше понимание основных событий, лежащих в основе переключения активности рук, а также формирования и распространения изгибов.
Подвижные реснички — самая ранняя из известных клеточных органелл , обнаруженная около 1647 года, когда ван Левенгук впервые увидел ресничные протисты в свой микроскоп. Хотя вопрос о механизме движения ресничек с того времени интересовал биологов и был предметом значительных спекуляций (Gray 1928), только с зарождением биологической электронной микроскопии в середине двадцатого века и появлением современной клеточной биологии несколько спустя годы появилась правдоподобная модель.Ключом к модели было понимание ультраструктуры ресничек. Исследования нескольких исследователей (Manton 1953, Fawcett and Porter 1959, Afzelius 1959, Gibbons IR и Grimstone 1960, Gibbons IR 1961) установили универсальность подвижной аксонемы ресничек 9 + 2 и ее важных субструктурных элементов, включая радиальные спицы и руки. , которые, как показали Гиббонс И.Р. и Роу (1965), являются цилиарными АТФазами, которые они назвали динеинами . Типичное изгибное движение ресничек и волнообразное движение биения жгутиков показано на рисунке 1.Рисунок был адаптирован на основе результатов количественного анализа прямого и обратного плавания (Brokaw and Kamiya, 1987). Chlamydomonas .
Хотя вопрос о механизме движения ресничек с того времени интересовал биологов и был предметом значительных спекуляций (Gray 1928), только с зарождением биологической электронной микроскопии в середине двадцатого века и появлением современной клеточной биологии несколько спустя годы появилась правдоподобная модель.Ключом к модели было понимание ультраструктуры ресничек. Исследования нескольких исследователей (Manton 1953, Fawcett and Porter 1959, Afzelius 1959, Gibbons IR и Grimstone 1960, Gibbons IR 1961) установили универсальность подвижной аксонемы ресничек 9 + 2 и ее важных субструктурных элементов, включая радиальные спицы и руки. , которые, как показали Гиббонс И.Р. и Роу (1965), являются цилиарными АТФазами, которые они назвали динеинами . Типичное изгибное движение ресничек и волнообразное движение биения жгутиков показано на рисунке 1.Рисунок был адаптирован на основе результатов количественного анализа прямого и обратного плавания (Brokaw and Kamiya, 1987). Chlamydomonas . На рис. 2 представлена ранняя электронная микрофотография рисунка 9 + 2 жаберных ресничек мидии, стандартного экспериментального организма для ранних исследований ресничек (Warner and Satir 1974), а также диаграмма, показывающая ось структуры 9 + 2 относительно плоскость изгиба.
На рис. 2 представлена ранняя электронная микрофотография рисунка 9 + 2 жаберных ресничек мидии, стандартного экспериментального организма для ранних исследований ресничек (Warner and Satir 1974), а также диаграмма, показывающая ось структуры 9 + 2 относительно плоскость изгиба.
Рис. 1.
Паттерны изгиба цилиарного (a) и жгутикового (b) движения у Chlamydomonas, зафиксированные с помощью высокоскоростной фотосъемки со вспышкой.Рисунки иллюстрируют изгибные паттерны движения, типичные для эпителиальных ресничек, и волнообразные паттерны движения, типичные для изгиба жгутиков эукариот. Серые стрелки указывают направление движения клеток. Черная полоса у основания ресничек или жгутиков имеет длину 5 микрометров и отмечает место прикрепления ресничек или жгутиков к клетке. Открытая стрелка на панели (а) указывает направление переднего изгиба ресничек. Открытая стрелка на панели (b) указывает направление распространения изгиба жгутика.Адаптировано с разрешения Brokaw и Kamiya (1987).
Рис. 1.
Паттерны изгиба цилиарного (a) и жгутикового (b) движения у Chlamydomonas, зафиксированные с помощью высокоскоростной фотосъемки со вспышкой. Рисунки иллюстрируют изгибные паттерны движения, типичные для эпителиальных ресничек, и волнообразные паттерны движения, типичные для изгиба жгутиков эукариот. Серые стрелки указывают направление движения клеток. Черная полоса у основания ресничек или жгутиков имеет длину 5 микрометров и отмечает место прикрепления ресничек или жгутиков к клетке.Открытая стрелка на панели (а) указывает направление переднего изгиба ресничек. Открытая стрелка на панели (b) указывает направление распространения изгиба жгутика. Адаптировано с разрешения Brokaw и Kamiya (1987).
Рис. 2.
Поперечный разрез подвижной реснички. (а) Электронная микрофотография единственной реснички из латеральных ресничек жабр моллюска при взгляде изнутри клетки по направлению к дистальному концу реснички. Дублет 1 расположен в положении «12 часов», а другие внешние микротрубочки дублета пронумерованы по часовой стрелке в направлении, в котором указывают плечи динеина. Также проиллюстрирован мост 5–6. Латеральные реснички бьются в плоскости, определяемой дублетом 1 и мостиком 5-6, и в этих ресничках эффективное или прямое направление изгиба направлено к мосту 5-6. Источник: перепечатано с разрешения Warner and Satir (1974). (b) Диаграмма, показывающая основные особенности аксонемы 9 + 2 из подвижных ресничек многоклеточных животных, включая микротрубочки наружного дублета и центральной пары, внешнее и внутреннее плечо динеина, выступы центральной пары, мостик 5–6, радиальные спицы, и регуляторный комплекс динеина и связи нексина.
Также проиллюстрирован мост 5–6. Латеральные реснички бьются в плоскости, определяемой дублетом 1 и мостиком 5-6, и в этих ресничках эффективное или прямое направление изгиба направлено к мосту 5-6. Источник: перепечатано с разрешения Warner and Satir (1974). (b) Диаграмма, показывающая основные особенности аксонемы 9 + 2 из подвижных ресничек многоклеточных животных, включая микротрубочки наружного дублета и центральной пары, внешнее и внутреннее плечо динеина, выступы центральной пары, мостик 5–6, радиальные спицы, и регуляторный комплекс динеина и связи нексина.
Рис. 2.
Поперечный разрез подвижной реснички. (а) Электронная микрофотография единственной реснички из латеральных ресничек жабр моллюска при взгляде изнутри клетки по направлению к дистальному концу реснички. Дублет 1 расположен в положении «12 часов», а другие внешние микротрубочки дублета пронумерованы по часовой стрелке в направлении, в котором указывают плечи динеина. Также проиллюстрирован мост 5–6. Латеральные реснички бьются в плоскости, определяемой дублетом 1 и мостиком 5-6, и в этих ресничках эффективное или прямое направление изгиба направлено к мосту 5-6. Источник: перепечатано с разрешения Warner and Satir (1974). (b) Диаграмма, показывающая основные особенности аксонемы 9 + 2 из подвижных ресничек многоклеточных животных, включая микротрубочки наружного дублета и центральной пары, внешнее и внутреннее плечо динеина, выступы центральной пары, мостик 5–6, радиальные спицы, и регуляторный комплекс динеина и связи нексина.
Источник: перепечатано с разрешения Warner and Satir (1974). (b) Диаграмма, показывающая основные особенности аксонемы 9 + 2 из подвижных ресничек многоклеточных животных, включая микротрубочки наружного дублета и центральной пары, внешнее и внутреннее плечо динеина, выступы центральной пары, мостик 5–6, радиальные спицы, и регуляторный комплекс динеина и связи нексина.
Форма ресничного биения хорошо изучена. Хотя иногда он может быть сильно изменчивым, даже на одной и той же клетке, типичный толчок ресничек обычно представляет собой короткий, очевидно жесткий эффективный ход, при котором ресничка указывает в направлении генерирования силы, и гибкий ход восстановления, при котором кончик ресничек медленнее отводится назад (рисунки 1а и 3), тогда как типичные сперматозоиды с 9 + 2 хвостами ресничек (называемые жгутиками ) бьют волнообразными движениями (рисунок 1b).И изгибное, и волнообразное движение (рисунок 1) вызывают поток жидкости, перемещение воды или слизи по реснитчатой поверхности (рисунок 1a) или продвижение реснитчатых клеток (рисунок 1b) через воду со скоростью примерно до 1 миллиметра в секунду, в зависимости от таких параметров, как длина ресничек и частота биений. Почти все реснички работают в гидродинамическом режиме с низким числом Рейнольдса, в котором силы вязкости имеют первостепенное значение, а силами инерции можно пренебречь (Holwill 1974). По сути, эффективный ход при изгибном движении (открытая стрелка на рисунке 1а) перемещается перпендикулярно поверхности ячейки, что вызывает скорость жидкости, примерно вдвое превышающую скорость хода восстановления, которая движется более параллельно поверхности.Гидромеханические аспекты движения ресничек были тщательно проанализированы и смоделированы (напр., Blake and Sleigh 1974, Smith DJ et al. 2007).
Почти все реснички работают в гидродинамическом режиме с низким числом Рейнольдса, в котором силы вязкости имеют первостепенное значение, а силами инерции можно пренебречь (Holwill 1974). По сути, эффективный ход при изгибном движении (открытая стрелка на рисунке 1а) перемещается перпендикулярно поверхности ячейки, что вызывает скорость жидкости, примерно вдвое превышающую скорость хода восстановления, которая движется более параллельно поверхности.Гидромеханические аспекты движения ресничек были тщательно проанализированы и смоделированы (напр., Blake and Sleigh 1974, Smith DJ et al. 2007).
Рисунок 3.
Диаграммы, иллюстрирующие (а) фазы изгиба ресничек и (б) геометрию скользящей модели микротрубочек для изгиба ресничек. Детали каждой диаграммы обсуждаются в тексте и иллюстрируют геометрию скольжения микротрубочек для эффективных и обратных изгибов микротрубочек, закрепленных в базальном теле и свободно скользящих на дистальной аксонеме.Модель основана на открытии того факта, что микротрубочки нерастяжимы и что их изгибы имеют форму дуг окружности. Источник: перепечатано с разрешения Satir (1968).
Источник: перепечатано с разрешения Satir (1968).
Рисунок 3.
Диаграммы, иллюстрирующие (а) фазы изгиба ресничек и (б) геометрию скользящей модели микротрубочек для изгиба ресничек. Детали каждой диаграммы обсуждаются в тексте и иллюстрируют геометрию скольжения микротрубочек для эффективных и обратных изгибов микротрубочек, закрепленных в базальном теле и свободно скользящих на дистальной аксонеме.Модель основана на открытии того факта, что микротрубочки нерастяжимы и что их изгибы имеют форму дуг окружности. Источник: перепечатано с разрешения Satir (1968).
В контексте 9 + 2 аксонемных механизмов нет выдающихся различий между органеллами, первоначально названными ресничками и теми, которые называются жгутиками . Ранние модели предполагали, что для возникновения инсульта девять дублетных микротрубочек модели 9 + 2 последовательно сокращаются, и это казалось биофизически правдоподобным, но с растущим пониманием того, что сокращение мышц зависит от скольжения актиновых и миозиновых нитей, люди начали считать, что сокращение ресничек также может быть вызвано скольжением. Какие элементы будут скользить? Когда Афзелиус (1959) впервые увидел руки, он предположил, что они могут вызывать скольжение дублетных микротрубочек.
Какие элементы будут скользить? Когда Афзелиус (1959) впервые увидел руки, он предположил, что они могут вызывать скольжение дублетных микротрубочек.
Satir (1963) подошел к проблеме, попытавшись использовать электронный микроскоп для фиксации структурных изменений внутри реснички по мере изменения формы биений. Биение можно было остановить, капнув раствор четырехокиси осмия на препарат, бьющийся метахронными волнами. Несколько исследователей, начиная с Gelei (1925, цитируется в Satir 1963), зафиксировали метахрональную волну на простейших, улавливая реснички в последовательных положениях биения вдоль длины волны.Сатир (1963, 1965) зафиксировал метахрональную волну латеральных ресничек мидии, впервые зафиксировав с помощью электронной микроскопии направленные в разные стороны реснички во время мазка. До фиксации частота биений ресничек составляла приблизительно 17 герц (Гц), поэтому биение занимало около 60 миллисекунд (мс). Тридцать одну индивидуальную ресничку измеряли на длине волны 11 микрометров (мкм) в фиксированном препарате, так что реснички были захвачены с разностью фаз около 2 мс между соседними ресничками (Satir 1967).
Сатир (1963, 1965) предположил, что после такой быстрой фиксации изучение тонкой структуры аксонемы ресничек, захваченных в разных положениях, покажет, как происходит сокращение. Что исследовать, было неочевидно, но затем Roth и Shigenaka (1964) показали, что в некоторых аксонемах кончики не сохраняют паттерн 9 + 2; некоторые из дуплетов стали синглетами. Можно представить себе, что если некоторые из дублетов сократятся, чтобы согнуть ресничку, скажем, в направлении эффективного удара, сокращенные дублеты станут синглетами и исчезнут с кончиков до того, как дублеты на противоположной стороне (на стороне восстановления). Инсульт).Затем, когда ресничка изгибалась в направлении восстановительного удара, был бы виден прямо противоположный результат: дублеты, которые сокращались и исчезали, были бы теми, которые были на стороне восстановительного удара, тогда как те, которые находились на стороне эффективного удара, сохранялись.
Чтобы проверить это, необходимо однозначно идентифицировать аксонемные дублеты в поперечном срезе кончика. К счастью, боковые реснички жабр имели особую перемычку между двумя дублетами (рис. 2), которые, казалось, находились в одном и том же положении в каждом поперечном сечении и которые, по-видимому, идентифицировали дублеты в направлении эффективного удара.Предполагалось, что мостик в этих ресничках был стабильным и что идентифицированные дублеты были дублетами 5 и 6 в соответствии с общепринятой нумерацией (фиг. 2). Еще одно предположение заключалось в том, что все дублеты имели одинаковую морфологическую длину в прямой ресничке до того, как произошло изгибание.
К счастью, боковые реснички жабр имели особую перемычку между двумя дублетами (рис. 2), которые, казалось, находились в одном и том же положении в каждом поперечном сечении и которые, по-видимому, идентифицировали дублеты в направлении эффективного удара.Предполагалось, что мостик в этих ресничках был стабильным и что идентифицированные дублеты были дублетами 5 и 6 в соответствии с общепринятой нумерацией (фиг. 2). Еще одно предположение заключалось в том, что все дублеты имели одинаковую морфологическую длину в прямой ресничке до того, как произошло изгибание.
Первоначальная демонстрация (Satir 1965) показала, что дублеты на концах ресничек эффективного наведения и на концах ресничек наведения на восстановление исчезали в разных последовательностях, причем дублеты 4–6 сохранялись в первых после дублетов 9, 1 и 2 (1 — это рядом с 9 в конце последовательности), а дублеты 9, 1 и 2 сохраняются в последнем после того, как 4–6 закончились (рис. 3).Этот результат не был совместим с простой сократительной моделью, как было заявлено, но фактически был полной противоположностью того, что ожидалось. Однако это можно объяснить, если для компенсации изгиба дуплеты скользят друг мимо друга, не меняя длины. Хотя это было спорным в то время, это стало оригинальным доказательством модели подвижности ресничек скользящих микротрубочек.
Однако это можно объяснить, если для компенсации изгиба дуплеты скользят друг мимо друга, не меняя длины. Хотя это было спорным в то время, это стало оригинальным доказательством модели подвижности ресничек скользящих микротрубочек.
\ begin {уравнение *} \ Delta l_n = d_n \, \ Sigma \ alpha, \ end {уравнение *}
, где d n — эффективный диаметр аксонемы для дублета n , а Σα — сумма изгибов ресничек, измеренная в радианах. Для дублетов 1 и 5, которые находятся на противоположных сторонах аксонемы, d n — это просто диаметр аксонемы, около 0,2 мкм, а значение Δ l , ожидаемое на кончике, составляет 3,5 нанометра (нм) на градус (°) изгиба. Изгиб, который образуется в начале эффективного хода, составляет около 100 °, что дает максимальное прогнозируемое смещение около 100 нм между соседними дублетами.
Для дублетов 1 и 5, которые находятся на противоположных сторонах аксонемы, d n — это просто диаметр аксонемы, около 0,2 мкм, а значение Δ l , ожидаемое на кончике, составляет 3,5 нанометра (нм) на градус (°) изгиба. Изгиб, который образуется в начале эффективного хода, составляет около 100 °, что дает максимальное прогнозируемое смещение около 100 нм между соседними дублетами. Уравнение может быть проверено последовательным разрезанием кончиков ресничек изогнутых ресничек в фиксированной метахрональной волне и непосредственным измерением Δ l .Каждая секция имела толщину примерно 100 нм. В пределах погрешности измерения, как для эффективных, так и для указывающих на восстановление ресничек, измеренное смещение кончика наконечника соответствовало предсказанному значению (рис. 3; Satir 1968). Более того, на концах было замечено несколько различных паттернов, которые соответствуют снимкам ресничек с разными полными изгибами вдоль аксонемы во время фиксированного удара. Паттерны кончиков и степень смещения дублетов были позже подтверждены разворачиванием изолированных аксонем ресничек Tetrahymena (Sale and Satir 1976).
Паттерны кончиков и степень смещения дублетов были позже подтверждены разворачиванием изолированных аксонем ресничек Tetrahymena (Sale and Satir 1976).
Главное возражение против этого анализа состоит в том, что фиксация может вызывать релаксацию сокращенных микротрубочек обратно до их исходной длины. Можно было бы возразить, что это маловероятно, потому что фиксация происходит очень быстро и потому что множественные изменения вдоль аксонемы — которые мы обсудим сейчас — несовместимы с расслаблением. Проблема была однозначно решена Саммерсом и Гиббонсом (1971), которые использовали микроскопию темного поля, чтобы напрямую продемонстрировать скольжение между внешними дублетными микротрубочками.
К началу 1970-х годов стало ясно, что если ресничную мембрану осторожно удалить с помощью мягкого моющего средства (такого как Triton X-100), образовавшаяся голая аксонема может быть реактивирована, чтобы победить добавлением аденозинтрифосфата (АТФ). . Гиббонс Б. Х. и Гиббонс (1972) показали, что для сперматозоидов морского ежа реактивированный ритм был идентичен биению живой спермы, что означало, что практически весь механизм образования и контроля изгиба находится в аксонеме, и этот вывод подтвердился во многих более поздних исследованиях. эксперименты.Мембрана поддерживала концентрацию АТФ и различных ионов на уровне, необходимом для подвижности. Он также обеспечивает такие сигналы, как цАМФ (циклический аденозинмонофосфат) или ионы кальция (Ca 2+ ), которые воздействуют на белки аксонемы, влияя на поведение ресничек.
эксперименты.Мембрана поддерживала концентрацию АТФ и различных ионов на уровне, необходимом для подвижности. Он также обеспечивает такие сигналы, как цАМФ (циклический аденозинмонофосфат) или ионы кальция (Ca 2+ ), которые воздействуют на белки аксонемы, влияя на поведение ресничек.
Саммерс и Гиббонс (1971) использовали этот препарат аксонем, но ненадолго переваривали их трипсином перед добавлением АТФ. Анализ просвечивающей электронной микроскопии показал, что целостность радиальной спицы была нарушена трипсином.Вместо того, чтобы реактивировать биение, эти аксонемы теперь раздвигались, в конечном итоге увеличиваясь до восьми или девяти раз длиннее исходной аксонемы. Этот результат подтвердил вывод о том, что основным взаимодействием в подвижности ресничек было скольжение дублета, приводимое в действие активностью динеиновой руки, так что руки на любом аксонемном дублете ( n ) производили скольжение соседнего дублета ( n + 1). Очевидно, динеиновые рукава могли активироваться по длине каждого дублета, и почти каждый дублет в аксонеме был способен скользить. Кроме того, считалось, что разница между скольжением и изгибом связана с целостностью радиальных спиц и междублетных звеньев.
Кроме того, считалось, что разница между скольжением и изгибом связана с целостностью радиальных спиц и междублетных звеньев.
Итак, внутри аксонемы есть две системы, необходимые для подвижности ресничек: (1) нечувствительная к трипсину система генерирования силы, основанная на взаимодействиях динеин-микротрубочка, способная генерировать неограниченное изотропное скольжение иногда более 10 мкм на дуплет без систематического изгиба. образование или распространение и (2) чувствительная к трипсину система управления, в зависимости от спица-центральная оболочка и других взаимодействий, где скользящий максимум около 0.1 мкм на дублет (т.е. между соседними дублетами) строго связан с систематическим образованием и распространением изгиба на высокой частоте (до 100 Гц). Brokaw (1989, 1991) предоставил прямое подтверждение и критический количественный анализ скольжения дублета во время образования изгиба и распространения в волнообразном движении. Относительное движение 40-нм золотых шариков, связанных с обнаженными внешними дублетными аксонемами жгутиков сперматозоидов во время АТФ-реактивированного плавания, согласуется со скользящей моделью взаимодействий дублетов. Вернон и Вулли (2002) смогли увидеть скольжение на кончике в соответствии с моделью точки переключения в сперме млекопитающих.
Вернон и Вулли (2002) смогли увидеть скольжение на кончике в соответствии с моделью точки переключения в сперме млекопитающих.
Форма волны и направление прогрессии изгиба являются эпифеноменами, зависящими от времени скольжения, переключения и управляющих событий в аксонеме и могут изменяться. Главной демонстрацией этого вывода является переход от плавательного изгибного движения вперед к волнообразному движению, ведущему к обратному плаванию и обратно у Chlamydomonas жгутиков, демонстрирующих реакцию избегания (сравните цилиарный, рисунок 1a, с движением жгутика, рисунок 1b).Это изменение может быть продемонстрировано на моделях клеток Chlamydomonas с демембранными, реактивированными АТФ или жгутиковых аппаратах путем добавления или удаления Ca 2+ из раствора для реактивации (Hyams and Borisy 1978, Kamiya and Witman 1984).
Сейл и Сатир (1977) расширили эксперименты Саммерса и Гиббонса (1971) с помощью анализа просвечивающей электронной микроскопии скользящих аксонем Tetrahymena . После скольжения можно было изучить перекрытие между соседними дублетами.Они обнаружили, что дублет n + 1 всегда располагался наверху дублета n в таких перекрытиях. Они пришли к выводу, что динеиновые рукава имели однородную полярность, всегда проходя по дуплету, к которому они были постоянно прикреплены, к основанию ресничек. С появлением систем для изучения полярности микротрубочек, цилиарное основание было определено как отрицательный конец дублета, а аксонемные динеины в совокупности, следовательно, были двигателем с отрицательным концом. Тесты изолированных внешних динеиновых плеч и многих компонентов внутренних динеиновых плеч в анализах in vitro подтвердили этот вывод (Vale and Toyoshima, 1988).
После скольжения можно было изучить перекрытие между соседними дублетами.Они обнаружили, что дублет n + 1 всегда располагался наверху дублета n в таких перекрытиях. Они пришли к выводу, что динеиновые рукава имели однородную полярность, всегда проходя по дуплету, к которому они были постоянно прикреплены, к основанию ресничек. С появлением систем для изучения полярности микротрубочек, цилиарное основание было определено как отрицательный конец дублета, а аксонемные динеины в совокупности, следовательно, были двигателем с отрицательным концом. Тесты изолированных внешних динеиновых плеч и многих компонентов внутренних динеиновых плеч в анализах in vitro подтвердили этот вывод (Vale and Toyoshima, 1988).
Следствие однородной полярности активного скольжения означает, что во время биения ресничек только некоторые дублеты или части дублетов имеют активные руки в любой момент времени. Когда дублет n + 1 движется к основанию дублета n , движение является пассивным, а руки дублета n в целом неактивны. Тщательное исследование ресничек жаберных мидий показало, что во время эффективного удара дублеты примерно на половине аксонемы вызывали активное скольжение, тогда как скольжение противоположной половины было пассивным, а во время восстановительного удара половинки менялись местами (Satir and Matsuoka 1989).Во время биения ресничек скользящая активность переключается между двумя половинами аксонемы, определяемыми линией, которая проходит между центральной парой и разделяет мост и дуплет 1 пополам (рис. 2). Эта гипотеза представляет собой модель точки переключения цилиарного биения (см. Также Morita and Shingyoji 2004, Hayashi and Shingyoji 2008).
Тщательное исследование ресничек жаберных мидий показало, что во время эффективного удара дублеты примерно на половине аксонемы вызывали активное скольжение, тогда как скольжение противоположной половины было пассивным, а во время восстановительного удара половинки менялись местами (Satir and Matsuoka 1989).Во время биения ресничек скользящая активность переключается между двумя половинами аксонемы, определяемыми линией, которая проходит между центральной парой и разделяет мост и дуплет 1 пополам (рис. 2). Эта гипотеза представляет собой модель точки переключения цилиарного биения (см. Также Morita and Shingyoji 2004, Hayashi and Shingyoji 2008).
Фиксация метахронной волны позволяет проследить развитие изгиба с разрешением электронного микроскопа. Если соединение спиц или междуплетов с центральной парой хотя бы частично отвечает за изгиб, как предполагали эксперименты Саммерса и Гиббонса (1971), скольжение должно вызывать смещение любой прикрепленной спицы или звена на величину скольжения. Когда изгиб происходит в основании ресничек, степень скольжения увеличивается по мере развития изгиба, и спицы наклоняются, но ни спицы, ни звенья не растягиваются непрерывно, поскольку скольжение продолжает увеличиваться; вместо этого, по мере роста изгиба вдоль аксонемы вовлекается больше спиц.
Когда изгиб происходит в основании ресничек, степень скольжения увеличивается по мере развития изгиба, и спицы наклоняются, но ни спицы, ни звенья не растягиваются непрерывно, поскольку скольжение продолжает увеличиваться; вместо этого, по мере роста изгиба вдоль аксонемы вовлекается больше спиц.
Warner и Satir (1974) рассмотрели, как наклон спицы может изменяться по мере роста изгиба аксонемы. В пределах изгиба по мере его роста все спицы каждой группы повторений 96-нм спиц наклоняются максимум на 33 ° от нормального.Поскольку углы спиц не лежат на радиусах, проходящих через центр кривизны изгиба, и поскольку скольжение накапливается в изгибе, спицы должны быть прикреплены к выступам оболочки вокруг центральной пары, когда происходит изгиб. Спицы выровнены с двумя рядами выступов вдоль каждой из центральных микротрубочек, которые образуют центральную оболочку. Выступы повторяются и образуют нониус с радиальными спицами в точном соотношении шести повторений выступов к одному повторению группы спиц. По мере развития базального изгиба количественно добавляются новые группы дистальных спиц.
По мере развития базального изгиба количественно добавляются новые группы дистальных спиц.
Наклон спицы во внешней дублетной микротрубочке с длиной повторяемости 96 нм остается постоянным по мере того, как изгиб увеличивается до максимальной кривизны, затем, когда изгиб проходит, спицы отсоединяются по мере выпрямления аксонемы. В прямых участках аксонемы, проксимальных или дистальных по отношению к изгибу, относительное положение групп спиц между любыми двумя дублетами остается постоянным по длине этой области, что означает, что скольжение происходит только в областях изгиба. В прямых участках аксонемы почти все спицы каждой группы нормальны (90 °) как по отношению к их исходному дублету, так и к центральным микротрубочкам.Функционально все эти спицы следует рассматривать как отделенные от выступов оболочки. Наблюдаемые радиальные конфигурации спиц строго подразумевают, что существует точный цикл отсоединения спиц-присоединения к центральной оболочке, который, как мы заключаем, составляет основную часть механизма преобразования активного междублетного скольжения в локальный изгиб.
Хотя изгибание требует активности dynein руки, локус активности руки вдоль аксонемы все еще неизвестен (см. Ниже и Lin et al. 2014).Может случиться так, что активность руки вдоль дуплета происходит только в положении сгибания. Также неизвестно, сколько циклов активности динеинового плеча необходимо для получения 96-нм единицы изгиба на одном дублете. In vitro , единственный динеин способен вызывать скольжение, но динеины внешнего плеча аксонемы имеют низкое соотношение фаза нагрузки: цикл, что означает, что для плавной непрерывной подвижности во время развития изгиба должны быть задействованы множественные динеины, действующие в противофазе. (В этой статье мы не останавливались на цикле генерации силы динеинов; отличные обзоры см. В Sakakibara and Oiwa 2011, Kikkawa 2013, and Roberts et al.2013). Разумное предположение состоит в том, что во время развития изгиба каждый динеин — как внутренний, так и внешний — в группе спиц на активном дублете потенциально циклически повторяется один раз.
Shingyoji и его коллеги (1977) провели эксперимент с освещением. Они прикрепили демембранную прямую аксонему морского ежа к микроигле и ионтофоретически добавили небольшое количество АТФ в середину аксонемы. Поскольку аксонема была прямой и ее концы были жесткими, при активации динеиновых плеч они образовывали равные и противоположные изгибы.Предположительно, согласно гипотезе точки переключения, активность вдоль половины аксонемы (дублеты 1–5; рисунок 2) в одной области уравновешена, противодействуя активности противоположной половины (дублеты 6–9 и 1; рисунок 2) дальше вдоль аксонемы. . Изгибание приводит к сопротивлению скольжению, а механическое напряжение, создаваемое действием руки на аксонему в одном положении, может передавать напряжение вдоль аксонемы, вероятно, через спицы и центральные парные крепления, чтобы активировать уравновешивающую активность руки в дублетах в более отдаленном положении — именно то, что необходим для волновой прогрессии в гипотезе точки переключения. Модель контроля кривизны Брокоу (1985) представляет собой упрощенный вид трансмиссии, который еще предстоит полностью уточнить. При нормальной перистальтике ресничек контроль такой передачи очень сложен, что позволяет большому количеству изгибных волн разной величины и времени распространяться по аксонеме. Более того, передача активности руки д. Происходить с довольно высокими скоростями, чтобы позволить завершить полный цикл биений вдоль аксонемы длиной 10-15 мкм менее чем за 10 миллисекунд в некоторых случаях.Дополнительные вопросы о колебании и изгибе ресничек обсуждаются в прекрасных недавних обзорах Brokaw (2009) и Lindemann and Lesich (2010).
Модель контроля кривизны Брокоу (1985) представляет собой упрощенный вид трансмиссии, который еще предстоит полностью уточнить. При нормальной перистальтике ресничек контроль такой передачи очень сложен, что позволяет большому количеству изгибных волн разной величины и времени распространяться по аксонеме. Более того, передача активности руки д. Происходить с довольно высокими скоростями, чтобы позволить завершить полный цикл биений вдоль аксонемы длиной 10-15 мкм менее чем за 10 миллисекунд в некоторых случаях.Дополнительные вопросы о колебании и изгибе ресничек обсуждаются в прекрасных недавних обзорах Brokaw (2009) и Lindemann and Lesich (2010).
Структурная и биохимическая сложность проливает свет на нерешенные проблемы модели точки переключения
Модель точки переключения была разработана в то время, когда разрешение электронного микроскопа и биохимия аксонем были в зачаточном состоянии. Предполагалось, что динеины внешнего и внутреннего плеча представляют собой зеркально отображаемые идентичные молекулы, что оболочка вокруг центральной пары функционально симметрична, так что микротрубочки центральной пары по существу идентичны, и что биохимия аксонем была по существу простой. Ни одно из этих предположений не подтвердилось. Чтобы расширить модель для учета подвижности, потребовались новые структурные, биохимические и генетические подходы.
Ни одно из этих предположений не подтвердилось. Чтобы расширить модель для учета подвижности, потребовались новые структурные, биохимические и генетические подходы.
Важно отметить, что асимметрия в организации центральных пар обязательно означает, что в любой момент цикла изгиба отдельные проекции центральных пар взаимодействуют с радиальными спицами из единственной уникальной внешней дублетной микротрубочки (Smith EF and Yang 2004). Следовательно, одна из моделей состоит в том, что центральная пара контролирует форму волны ресничек путем избирательной регуляции активности динеина в любой точке цикла биений на единственной внешней дублетной микротрубочке или их подмножестве.Более того, помимо асимметрии аппарата центральной пары, общей для подвижных ресничек, у многих организмов, включая Chlamydomonas , центральная пара вращается по мере изгиба ресничек (Smith EF and Yang 2004). Следовательно, в этих случаях центральная пара рассматривается как распределитель, и при вращении во время цикла биений ресничек центральная пара, как полагают, систематически взаимодействует с разными радиальными спицами, выступающими из разных внешних дублетных микротрубочек. Предположительно, это приводит к систематической активации динеиновых моторов на разных фазах биения.Однако точная роль вращения центральной пары в ресничках у некоторых организмов не решена (Mitchell and Nakatsugawa 2004).
Предположительно, это приводит к систематической активации динеиновых моторов на разных фазах биения.Однако точная роль вращения центральной пары в ресничках у некоторых организмов не решена (Mitchell and Nakatsugawa 2004).
Асимметрия в аппарате центральной пары также подразумевает роль в избирательном контроле прямого и обратного изгиба и точки переключения, описанной выше и необходимой для чередующихся изгибов ресничек. Например, при сравнении движения жгутиков Chlamydomonas дикого типа и мутанта , Brokaw с коллегами (1982) определили, что центральная пара-радиальная система спиц важна для преобразования симметричного прямого и обратного изгибов в асимметричные изгибы, необходимые для эффективное сгибательное движение ресничек и нормальные физиологические силы.Более конкретно, одна из функций радиальных спиц, по-видимому, заключается в относительном подавлении развития обратного изгиба по сравнению с кривизной прямого изгиба (Brokaw et al. 1982).
Мутация hydin у Chlamydomonas и у мышей выявила роль системы центральная пара-радиальная спица в контроле переключения между прямым и обратным изгибами (Lechtreck and Witman 2007, Lechtreck et al. 2008). Примечательно, что мутация в гене HYDIN приводит к большому дефекту изгиба ресничек, но структурный анализ выявляет только очень маленький дефект в структуре аксонемы.И у Chlamydomonas , и у мышей проекция единственной центральной пары является единственным отсутствующим структурным компонентом мутантных аксонем. И у Chlamydomonas , и у мыши мутация hydin приводит к относительной нерешительности или срыву между прямым и обратным поворотами. Следовательно, самая простая интерпретация состоит в том, что мутация hydin влияет на механизм точки переключения, необходимый для переключения между прямым и обратным изгибами (см. Комментарий Smith EF 2007).В совокупности данные показывают, что центральная пара — система радиальных спиц играет центральную роль в переключении между прямым и обратным изгибами и в управлении ими — то есть, какие дублеты имеют активные плечи, когда изгиб формируется и прогрессирует, — и в точном контроле кривизна в каждом изгибе.
2008). Примечательно, что мутация в гене HYDIN приводит к большому дефекту изгиба ресничек, но структурный анализ выявляет только очень маленький дефект в структуре аксонемы.И у Chlamydomonas , и у мышей проекция единственной центральной пары является единственным отсутствующим структурным компонентом мутантных аксонем. И у Chlamydomonas , и у мыши мутация hydin приводит к относительной нерешительности или срыву между прямым и обратным поворотами. Следовательно, самая простая интерпретация состоит в том, что мутация hydin влияет на механизм точки переключения, необходимый для переключения между прямым и обратным изгибами (см. Комментарий Smith EF 2007).В совокупности данные показывают, что центральная пара — система радиальных спиц играет центральную роль в переключении между прямым и обратным изгибами и в управлении ими — то есть, какие дублеты имеют активные плечи, когда изгиб формируется и прогрессирует, — и в точном контроле кривизна в каждом изгибе. Однако эти модели требуют дальнейшего прямого тестирования.
Однако эти модели требуют дальнейшего прямого тестирования.
Мутанты Chlamydomonas были особенно информативными, давая понимание состава и структуры аксонемы, регуляции подвижности, роли различных двигателей динеина, а также сборки и регуляции длины ресничек (Avasthi and Marshall 2012).Здесь мы сосредотачиваемся на том, как Chlamydomonas способствовало нашему пониманию механизма и регуляции изгиба ресничек и, в частности, как оно способствовало знанию общих ролей внешнего и внутреннего динеина в контроле частоты сердечных сокращений и формы волны. .
Первоначально и чаще всего аксонемные динеины называются внешними и внутренними динеиновыми ветвями, но в действительности существует множество различных консервативных динеиновых моторов, которые сильно локализованы в аксонеме и служат особым целям для контроля над ней. движение (Кинг и Камия 2009).Наиболее изученным аксонемным динеином является внешнее плечо динеина. Внешние динеиновые плечи очень сложны с биохимической точки зрения: каждое состоит из двух или трех различных динеиновых АТФаз и по крайней мере 16 различных субъединиц. Они собраны индивидуально в ряды на дублетах с номерами 2–9 в аксонемах Chlamydomonas и повторяются с регулярным периодом 24 нм вдоль каждого внешнего дублета (см. Рисунок 4; Гуденаф и Хойзер, 1982). Вдоль большей части аксонемы каждое внешнее плечо динеина структурно — и, обычно, биохимически — идентично своему соседу.Мутации в генах, которые кодируют структурные белки внешнего плеча динеина или связанные факторы, участвующие в нацеливании и сборке внешнего плеча динеина, могут привести к неспособности собрать всю или часть внешнего плеча динеина и, как следствие, к нарушению подвижности (King и Камия 2009).
движение (Кинг и Камия 2009).Наиболее изученным аксонемным динеином является внешнее плечо динеина. Внешние динеиновые плечи очень сложны с биохимической точки зрения: каждое состоит из двух или трех различных динеиновых АТФаз и по крайней мере 16 различных субъединиц. Они собраны индивидуально в ряды на дублетах с номерами 2–9 в аксонемах Chlamydomonas и повторяются с регулярным периодом 24 нм вдоль каждого внешнего дублета (см. Рисунок 4; Гуденаф и Хойзер, 1982). Вдоль большей части аксонемы каждое внешнее плечо динеина структурно — и, обычно, биохимически — идентично своему соседу.Мутации в генах, которые кодируют структурные белки внешнего плеча динеина или связанные факторы, участвующие в нацеливании и сборке внешнего плеча динеина, могут привести к неспособности собрать всю или часть внешнего плеча динеина и, как следствие, к нарушению подвижности (King и Камия 2009).
Рисунок 4.
Ультраструктура аксонемы Chlamydomonas. Срезы криоэлектронной томографии показывают (a) продольный, (b) трехмерный вид и (c) поперечный разрез аксонемы Chlamydomonas. Красные прямоугольники выделяют одну 96-нанометровую (нм) аксонемную повторяющуюся единицу на каждом изображении. (d, e) Визуализация изоповерхности и (f, g) упрощенная схема показывают усредненный аксонемный повтор 96 нм в (d, f) продольной и (e, g) поперечной ориентации. Поперечный срез берут рядом с радиальной спицей 2, если смотреть от проксимального до дистального конца. Выделены ключевые структуры аксонемы: A- и B-канальцы (A t , B t ), регуляторный комплекс нексин-динеин (N-DRC), радиальные спицы (RS1, RS2), кальмодулин и связанные спицы. комплекс (CSC), а также внутренние и внешние динеиновые рукава (IA, OA).Динеины внутреннего плеча включают комплекс I1 (динеин f α и β) и динеины a – g. Источник: адаптировано с разрешения Heuser и коллег (2012a, 2012b).
Красные прямоугольники выделяют одну 96-нанометровую (нм) аксонемную повторяющуюся единицу на каждом изображении. (d, e) Визуализация изоповерхности и (f, g) упрощенная схема показывают усредненный аксонемный повтор 96 нм в (d, f) продольной и (e, g) поперечной ориентации. Поперечный срез берут рядом с радиальной спицей 2, если смотреть от проксимального до дистального конца. Выделены ключевые структуры аксонемы: A- и B-канальцы (A t , B t ), регуляторный комплекс нексин-динеин (N-DRC), радиальные спицы (RS1, RS2), кальмодулин и связанные спицы. комплекс (CSC), а также внутренние и внешние динеиновые рукава (IA, OA).Динеины внутреннего плеча включают комплекс I1 (динеин f α и β) и динеины a – g. Источник: адаптировано с разрешения Heuser и коллег (2012a, 2012b).
Рис. 4.
Ультраструктура аксонемы Chlamydomonas. Срезы криоэлектронной томографии показывают (a) продольный, (b) трехмерный вид и (c) поперечный разрез аксонемы Chlamydomonas. Красные прямоугольники выделяют одну 96-нанометровую (нм) аксонемную повторяющуюся единицу на каждом изображении. (d, e) Визуализация изоповерхности и (f, g) упрощенная схема показывают усредненный аксонемный повтор 96 нм в (d, f) продольной и (e, g) поперечной ориентации.Поперечный срез берут рядом с радиальной спицей 2, если смотреть от проксимального до дистального конца. Выделены ключевые структуры аксонемы: A- и B-канальцы (A t , B t ), регуляторный комплекс нексин-динеин (N-DRC), радиальные спицы (RS1, RS2), кальмодулин и связанные спицы. комплекс (CSC), а также внутренние и внешние динеиновые рукава (IA, OA). Динеины внутреннего плеча включают комплекс I1 (динеин f α и β) и динеины a – g. Источник: адаптировано с разрешения Heuser и коллег (2012a, 2012b).
(d, e) Визуализация изоповерхности и (f, g) упрощенная схема показывают усредненный аксонемный повтор 96 нм в (d, f) продольной и (e, g) поперечной ориентации.Поперечный срез берут рядом с радиальной спицей 2, если смотреть от проксимального до дистального конца. Выделены ключевые структуры аксонемы: A- и B-канальцы (A t , B t ), регуляторный комплекс нексин-динеин (N-DRC), радиальные спицы (RS1, RS2), кальмодулин и связанные спицы. комплекс (CSC), а также внутренние и внешние динеиновые рукава (IA, OA). Динеины внутреннего плеча включают комплекс I1 (динеин f α и β) и динеины a – g. Источник: адаптировано с разрешения Heuser и коллег (2012a, 2012b).
В соответствии с новаторскими исследованиями Гиббонса Б.Х. и Гиббонса (1973), наиболее заметным последствием сбоя сборки внешнего плеча динеина является снижение частоты биений ресничек (Brokaw and Kamiya 1987, Brokaw 1994, Kamiya 2002). Внешнее плечо динеина и частота биений ресничек могут регулироваться фосфорилированием и изменениями кальция (Christensen et al. 2001, King and Kamiya 2009, King 2010). У инфузорий Paramecium и Tetrahymena увеличение цАМФ вокруг аксонемы вызывает более быстрое плавание за счет фосфорилирования небольшого белка, связанного с внешним динеиновым плечом (Christensen et al.2001). In vitro , фосфорилированное внешнее плечо динеина вызывает более быстрое скольжение микротрубочек, что означает, что меньше времени требуется для того, чтобы произвести такое же количество скольжения в течение цикла биений, то есть частота биений выше. Разнообразные доказательства также указывают на то, что внешние динеиновые рукава реагируют на механическое возмущение аксонемы (Hayashibe et al. 1997). Механическая обратная связь, вероятно, важна для переключения между активным и неактивным состояниями и необходима для прямого и обратного изгиба.В соответствии с управлением с механической обратной связью, как обсуждалось выше (Shingyoji et al. 1977), изгиб аксонемы может активировать управляемое динеином скольжение микротрубочек (Morita and Shingyoji 2004, Hayashi and Shingyoji 2008).
2001, King and Kamiya 2009, King 2010). У инфузорий Paramecium и Tetrahymena увеличение цАМФ вокруг аксонемы вызывает более быстрое плавание за счет фосфорилирования небольшого белка, связанного с внешним динеиновым плечом (Christensen et al.2001). In vitro , фосфорилированное внешнее плечо динеина вызывает более быстрое скольжение микротрубочек, что означает, что меньше времени требуется для того, чтобы произвести такое же количество скольжения в течение цикла биений, то есть частота биений выше. Разнообразные доказательства также указывают на то, что внешние динеиновые рукава реагируют на механическое возмущение аксонемы (Hayashibe et al. 1997). Механическая обратная связь, вероятно, важна для переключения между активным и неактивным состояниями и необходима для прямого и обратного изгиба.В соответствии с управлением с механической обратной связью, как обсуждалось выше (Shingyoji et al. 1977), изгиб аксонемы может активировать управляемое динеином скольжение микротрубочек (Morita and Shingyoji 2004, Hayashi and Shingyoji 2008). Кроме того, эти данные могут также косвенно относиться к важной модели механического контроля изгиба, называемой геометрической гипотезой сцепления , которая предусматривает изменения междублетных расстояний, соответствующих активности руки и производству изгибов, связанных с искажением аксонемы во время изгиба (для полное обсуждение см. Lindemann 2011).Предполагаемое искажение аксонемы во время изгиба наблюдали с помощью электронной микроскопии (Lindemann and Mitchell 2007).
Кроме того, эти данные могут также косвенно относиться к важной модели механического контроля изгиба, называемой геометрической гипотезой сцепления , которая предусматривает изменения междублетных расстояний, соответствующих активности руки и производству изгибов, связанных с искажением аксонемы во время изгиба (для полное обсуждение см. Lindemann 2011).Предполагаемое искажение аксонемы во время изгиба наблюдали с помощью электронной микроскопии (Lindemann and Mitchell 2007).
Внутренние рукава динеина намного сложнее, чем внешние рукава динеина (обзоры см. В Kamiya 2002, King and Kamiya 2009). Вдоль аксонемного 96-нм повтора на дублетной микротрубочке (рисунок 4, который имеет четыре идентичных внешних плеча динеина) внутренние плечи динеина включают по крайней мере семь различных динеинов, которые различаются по составу и расположению (см. Bui et al.2012). Внутренние плечи динеина были охарактеризованы посредством биохимического фракционирования компонентов динеина и с помощью электронной микроскопии аксонем, отсутствующих подмножества внутренних плеч динеина (King and Kamiya 2009). В аксонемах Chlamydomonas , у которых отсутствует подмножество внутренних динеиновых плеч, изменяется форма волны цилиарной системы, фототаксис нарушается и скорость плавания клеток замедляется. Прямой анализ биения ресничек с помощью высокоскоростной видеозаписи подтвердил, что нарушение сборки любого из отдельных внутренних динеиновых плеч приводит к изменению формы волны ресничек, параметра, который важен для эффективного биения ресничек и физиологии (Brokaw and Kamiya 1987). .
В аксонемах Chlamydomonas , у которых отсутствует подмножество внутренних динеиновых плеч, изменяется форма волны цилиарной системы, фототаксис нарушается и скорость плавания клеток замедляется. Прямой анализ биения ресничек с помощью высокоскоростной видеозаписи подтвердил, что нарушение сборки любого из отдельных внутренних динеиновых плеч приводит к изменению формы волны ресничек, параметра, который важен для эффективного биения ресничек и физиологии (Brokaw and Kamiya 1987). .
Точная роль каждого внутреннего плеча динеина еще не изучена. Однако одно из двуглавых внутренних плечей динеина, называемое динеином I1 , считается особенно важным для контроля изгиба аксонемы. Активность I1 динеина регулируется киназами и фосфатазами, локализованными в аксонеме (см. Обзор Wirschell et al. 2011). Изменения активности I1 можно измерить по изменениям скорости скольжения микротрубочек, но пока не известно, как изменения скорости соответствуют изменениям в изгибе.Одна возможность состоит в том, что увеличение скорости скольжения, производимой внутренними плечами динеина без изменения частоты биений, будет соответствовать увеличению величины изгиба. Кроме того, I1 динеин может регулировать изгиб посредством контроля активности др. Динеинов, включая внешние динеиновые плечи и одноголовые внутренние динеиновые плечи (Kotani et al. 2007, Yamamoto et al. 2013).
Кроме того, I1 динеин может регулировать изгиб посредством контроля активности др. Динеинов, включая внешние динеиновые плечи и одноголовые внутренние динеиновые плечи (Kotani et al. 2007, Yamamoto et al. 2013).
В целом, мы еще не знаем функцию каждого из аксонемных одноголовых динеинов. Однако анализ мутанта под названием ida9 показал, что одно из внутренних плеч динеина, динеин c, необходимо для движения ресничек в вязкой жидкости (Yagi et al.2005). Кроме того, мощный скрининг выявил новые мутанты, которые регулируют активность внутреннего плеча динеина (Kamiya et al. 1991, Kamiya 2002). Напр., Скрининг выявил ферменты, ответственные за полиглутамилирование, и что эта посттрансляционная модификация тубулина является критической для активности в подмножестве внутренних плеч динеина (Kubo et al. 2010).
Центральная пара и радиальные спицы необходимы для нормальной перистальтики ресничек и управления двигателем динеина
У Chlamydomonas нарушение сборки центральной пары или радиальных спиц приводит либо к цилиарному параличу (Witman et al. 1978, Smith EF and Yang 2004) или в сильно измененных и непродуктивных сгибательных движениях. Вместе аппарат центральной пары и радиальные спицы действуют с помощью как механической, так и химической передачи сигналов, чтобы в конечном итоге контролировать активность аксонемального динеина (Smith EF and Yang 2004). Механизм взаимодействия между центральной парой и радиальной спицей был подтвержден в недавних экспериментальных исследованиях с участием Chlamydomonas (Oda et al. 2014). Они показали, что добавление неспецифических белков к головке лучевой спицы могло подавить паралич парализованного мутанта центральной пары, у которого отсутствовала часть выступов центральной пары.Простейшая интерпретация этого результата состоит в том, что добавленные белки обеспечивали физическое взаимодействие между головкой спицы и выступами, которое требуется для активации аксонемных динеинов.
1978, Smith EF and Yang 2004) или в сильно измененных и непродуктивных сгибательных движениях. Вместе аппарат центральной пары и радиальные спицы действуют с помощью как механической, так и химической передачи сигналов, чтобы в конечном итоге контролировать активность аксонемального динеина (Smith EF and Yang 2004). Механизм взаимодействия между центральной парой и радиальной спицей был подтвержден в недавних экспериментальных исследованиях с участием Chlamydomonas (Oda et al. 2014). Они показали, что добавление неспецифических белков к головке лучевой спицы могло подавить паралич парализованного мутанта центральной пары, у которого отсутствовала часть выступов центральной пары.Простейшая интерпретация этого результата состоит в том, что добавленные белки обеспечивали физическое взаимодействие между головкой спицы и выступами, которое требуется для активации аксонемных динеинов.
Передача сигналов от центральной пары и радиальных спиц к двигателям dynein
Анализ мутантов Chlamydomonas также выявил консервативные компоненты аксонемы, которые передают сигналы от центральной пары и структур радиальных спиц к динеиновым моторам. Наиболее примечательными являются комплекс, связанный с кальмодулином и спицами (CSC; Dymek et al. 2011, Heuser et al. 2012a) и DRC; см. следующий раздел для получения информации из недавних анализов криоэлектронной томографии (крио-ET) аксонемы (обзоры см. Heuser et al. 2009, Porter 2012) и комплекса динеина Mia-I1 (Yamamoto et al. 2013). CSC и DRC играют роль в управлении двигателями динеина: они оба идеально расположены для соединения радиальных спиц с внешними дублетами и двигателями динеина.В настоящее время важной целью является определение того, как кальций и комплексы кальмодулина, расположенные в центральном аппарате и CSC, управляют аксонемными динеинами. Динеин I1 и связанный с ним комплекс Mia (Yamamoto et al. 2013) также могут играть роль, сходную с ролью DRC, в регуляции или устойчивости к управляемому динеином скольжению микротрубочек и в контроле изгиба аксонемы.
Наиболее примечательными являются комплекс, связанный с кальмодулином и спицами (CSC; Dymek et al. 2011, Heuser et al. 2012a) и DRC; см. следующий раздел для получения информации из недавних анализов криоэлектронной томографии (крио-ET) аксонемы (обзоры см. Heuser et al. 2009, Porter 2012) и комплекса динеина Mia-I1 (Yamamoto et al. 2013). CSC и DRC играют роль в управлении двигателями динеина: они оба идеально расположены для соединения радиальных спиц с внешними дублетами и двигателями динеина.В настоящее время важной целью является определение того, как кальций и комплексы кальмодулина, расположенные в центральном аппарате и CSC, управляют аксонемными динеинами. Динеин I1 и связанный с ним комплекс Mia (Yamamoto et al. 2013) также могут играть роль, сходную с ролью DRC, в регуляции или устойчивости к управляемому динеином скольжению микротрубочек и в контроле изгиба аксонемы.
Супрессорные мутации в
Chlamydomonas и DRC Основное открытие для понимания контроля аксонемных динеинов стало результатом классических генетических исследований Huang et al. (1982).Генетический скрининг выявил новые гены, которые подавляли паралич (то есть спасательную подвижность) у мутантов с радиальными спицами или центральной парой, так что мутанты снова становились подвижными. Супрессорные мутанты включали новые мутации в моторных тяжелых цепях динеина, которые восстанавливали движение без восстановления радиальной спицы и без дефектов центральной пары. Эти результаты показывают, что в отсутствие радиальных спиц и центральной пары моторная активность динеина подавляется по всей аксонеме, но она может быть восстановлена любым молекулярным изменением, которое позволяет динеину становиться активным без участия пары спица-центральная часть. система.
(1982).Генетический скрининг выявил новые гены, которые подавляли паралич (то есть спасательную подвижность) у мутантов с радиальными спицами или центральной парой, так что мутанты снова становились подвижными. Супрессорные мутанты включали новые мутации в моторных тяжелых цепях динеина, которые восстанавливали движение без восстановления радиальной спицы и без дефектов центральной пары. Эти результаты показывают, что в отсутствие радиальных спиц и центральной пары моторная активность динеина подавляется по всей аксонеме, но она может быть восстановлена любым молекулярным изменением, которое позволяет динеину становиться активным без участия пары спица-центральная часть. система.
Тот же самый скрининг генетических супрессоров (Huang et al. 1982) выявил другой регуляторный комплекс, который Piperno в 1994 назвал DRC. Последующие исследования дополнительно определили белковые субъединицы DRC (для обзора см. Porter 2012). Важным недавним достижением, которое стало возможным благодаря увеличенному разрешению крио-ET, является открытие того, что DRC также является междублетным звеном нексина. Поэтому структура DRC теперь называется регуляторным комплексом нексин-динеин (N-DRC; Heuser et al.2009 г.). Недавние исследования также показали биохимически-структурное взаимодействие между N-DRC и промежуточными цепями внешнего плеча динеина, физически связывающими внешнее плечо динеина и N-DRC (Oda et al. 2013). Следовательно, N-DRC, по-видимому, играет роль в регуляции моторов динеина и служит междублетным звеном, так что он, вероятно, выравнивает внешние дублеты для обеспечения эффективных взаимодействий между моторами динеина и B-канальцем соседней микротрубочки дублета (Bower и др., 2013).Чтобы приспособиться к скольжению дублетных микротрубочек, все междублетные связи N-DRC-внешнее плечо динеина на скользящем дублете должны в какой-то момент разрывать их связь с соседней B-микротрубочкой (Holwill and Satir 1990). В окончательном понимании модели точки переключения будет важно знать способ, которым регулируются связи N-DRC, и время разрыва и реформации для активных и пассивно скользящих половин аксонемы.
Поэтому структура DRC теперь называется регуляторным комплексом нексин-динеин (N-DRC; Heuser et al.2009 г.). Недавние исследования также показали биохимически-структурное взаимодействие между N-DRC и промежуточными цепями внешнего плеча динеина, физически связывающими внешнее плечо динеина и N-DRC (Oda et al. 2013). Следовательно, N-DRC, по-видимому, играет роль в регуляции моторов динеина и служит междублетным звеном, так что он, вероятно, выравнивает внешние дублеты для обеспечения эффективных взаимодействий между моторами динеина и B-канальцем соседней микротрубочки дублета (Bower и др., 2013).Чтобы приспособиться к скольжению дублетных микротрубочек, все междублетные связи N-DRC-внешнее плечо динеина на скользящем дублете должны в какой-то момент разрывать их связь с соседней B-микротрубочкой (Holwill and Satir 1990). В окончательном понимании модели точки переключения будет важно знать способ, которым регулируются связи N-DRC, и время разрыва и реформации для активных и пассивно скользящих половин аксонемы.
Анализ крио-ET
Новый уровень разрешения аксонемы был достигнут с помощью крио-ЭТ.Эти данные уже оказали большое влияние на понимание движения ресничек. Была рассмотрена базовая методология (например, Nicastro 2009, Bui and Ishikawa 2013) и описаны методы, которые теперь объединяют структурную локализацию с крио-ET (Oda and Kikkawa 2013). Вкратце, процесс включает быстрое замораживание живых клеток или изолированных аксонем с последующей электронной микроскопией с низкой дозой облучения структур в замороженном нативном состоянии, сбор данных, компьютерную томографию и усреднение изображений, которые позволяют получать изображения с высоким разрешением в три измерения.В сочетании с очень информативными структурными мутантами в Chlamydomonas появилась картина аксонемной структуры с очень высоким разрешением. Новая структурная информация обобщена на рисунке 4, включая примеры электронных томограмм (рисунки 4a-4c), усредненные значения и трехмерные изображения изоповерхности (рисунки 4d, 4e), а также сводные схематические диаграммы одиночной дублетной микротрубочки в продольном и поперечном сечениях (рисунок 4е, 4ж).
Подход крио-ET предлагает множество преимуществ: высокое разрешение, приближение и превышение 3 нм, выявление задержанной субструктуры каждого аксонемного компонента; первозданная сохранность за счет быстрого замораживания; и 3D визуализация структур в интактных органеллах.Примечательные особенности на рисунке 4 включают определение продольного 96-нм аксонемного повтора (рисунки 4 D и E), который, вероятно, является основной единицей активности вдоль аксонемы и субструктуры внешних плеч динеина, включая разрешение глобулярные моторные домены (рис. 4d – 4g). Также показаны расположение и субструктура каждого внутреннего плеча динеина (I1 / f динеин и динеины a – g; Bui et al. 2012, Heuser et al. 2012b, Lin et al. 2014), N-DRC (Heuser et al. 2009), CSC (Dymek et al.2011 г., Heuser et al. 2012a) и радиальные спицы (Pigino et al. 2011, Barber et al. 2012, Oda et al. 2014). Кроме того, крио-ЭТ выявила субструктуру аппарата центральной пары (Carbajal-Gonzalez et al. 2013, Oda et al. 2014).
2014).
За относительно короткое время эти структурные достижения внесли новое понимание структурных основ изгиба ресничек. Например, крио-ЭТ также выявил физические связи между внешними плечами динеина, внутренними плечами динеина и N-DRC, которые могут объяснить координацию активности между структурами (например,г., Oda et al. 2013). Этот результат начинает отвечать на главный вопрос — вопрос о том, как координируется активность внешних и внутренних рукавов динеина (Kamiya 2002). Наряду с другими биофизическими и структурными исследованиями крио-ЭТ также выявило структурную основу силового удара аксонемных динеинов (Lin et al. 2014). Обещание этих новых исследований состоит в том, что эти структурные достижения будут определять структурные изменения, связанные с изгибом ресничек, и будут непосредственно тестировать модели для формирования изгиба ресничек и распространения изгиба.Например, структурный анализ быстро замороженных аксонем живых сперматозоидов морского ежа показывает, что структурное состояние динеиновых моторов на дублете, расположенном на внутренней стороне изгиба, отличается от состояния динеинов, расположенных на дублетах на противоположной стороне отгиба. axoneme, на внешней стороне изгиба (см. дополнительную фигуру 1 у Lin et al. 2014). Эти данные, по-видимому, подтверждают модель точки переключения для изгиба аксонемы, возможно, раскрывая «включенные» и «выключенные» состояния динеинов (см. Brokaw, 2009).Как и ожидалось, использование крио-ЭТ и консервация посредством замораживания, дальнейший анализ аксонем сперматозоидов морского ежа, мутантов Chlamydomonas и ресничек, сохраненных на стадиях метахрональных биений, обеспечит исчерпывающую картину структурных изменений аксонемы и динеина, связанных с инициированием изгиба и распространение изгиба.
axoneme, на внешней стороне изгиба (см. дополнительную фигуру 1 у Lin et al. 2014). Эти данные, по-видимому, подтверждают модель точки переключения для изгиба аксонемы, возможно, раскрывая «включенные» и «выключенные» состояния динеинов (см. Brokaw, 2009).Как и ожидалось, использование крио-ЭТ и консервация посредством замораживания, дальнейший анализ аксонем сперматозоидов морского ежа, мутантов Chlamydomonas и ресничек, сохраненных на стадиях метахрональных биений, обеспечит исчерпывающую картину структурных изменений аксонемы и динеина, связанных с инициированием изгиба и распространение изгиба.
Выводы
Мы стоим на новом этапе понимания движения ресничек в молекулярных деталях, которые вряд ли представлялись возможными, когда были впервые предложены скользящая модель и гипотеза точки переключения.В целом основная гипотеза, кажется, выдержала проверку временем. Есть еще важные базовые вопросы, которые необходимо решить, например, как динамические связи между структурами внутри аксонемы соответствуют точному развитию и прогрессированию изгиба (Brokaw 2009). Новые возможности включают определение функции посттрансляционных модификаций тубулина для регуляции активности динеина (например, Kubo et al. 2010). Благодаря сочетанию новых генетических и структурных методов, как было проиллюстрировано изобретательным использованием мутантов Chlamydomonas и крио-ET (e.g., Oda et al., 2014), которые привели к новому пониманию сложности механизма моторики, мы можем дать содержательный ответ на многие из оставшихся вопросов.
Благодарности
Мы благодарны Алекса Маттейсес за помощь с цифрами. Мы также благодарны всем людям, с которыми мы работали все эти годы, нашим наставникам, студентам и нашим коллегам. Эта статья была поддержана грантом WSS от Национальных институтов здравоохранения.
Цитированные источники
.Электронная микроскопия хвоста сперматозоида; результаты, полученные с новым фиксатором
,Journal of Biophysical and Biochemical Cytology
,1959
, vol.5
(стр.269
—278
),.Этапы цилиогенеза и регуляции длины ресничек
,Дифференциация
,2012
, т.83
(стр.S30
—S42
),,,,.Трехмерная структура радиальных спиц выявляет неоднородность и взаимодействие с динеинами в жгутиках Chlamydomonas.
,Molecular Biology of the Cell
,2012
, vol.23
(стр.111
—120
),.Механика цилиарного движения
,Биологические обзоры Кембриджского философского общества
,1974
, vol.49
(стр.85
—125
),,,,,,,.N-DRC образует консервативный биохимический комплекс, который поддерживает внешнее выравнивание дублетов и ограничивает скольжение микротрубочек в подвижных аксонемах
,Molecular Biology of the Cell
,2013
, vol.24
(стр.1134
—1152
).Несинусоидальные изгибные волны жгутиков сперматозоидов
,Journal of Experimental Biology
,1965
, vol.43
(стр.155
—169
).Компьютерное моделирование движения жгутиков: VI. Простые модели с контролируемой кривизной определены не полностью
,Biophysical Journal
,1985
, vol.48
(стр.633
—642
).Прямые измерения скольжения между внешними дублетными микротрубочками в плавающих жгутиках сперматозоидов
,Science
,1989
, vol.243
(стр.1593
—1596
).Скольжение микротрубочек в плавающих жгутиках сперматозоидов: прямые и косвенные измерения на морских ежах и оболочках сперматозоидов
,Journal of Cell Biology
,1991
, vol.114
(стр.1201
—1215
).Контроль изгиба жгутиков: новая повестка дня, основанная на разнообразии динеина
,Подвижность клеток и цитоскелет
,1994
, vol.28
(стр.199
—204
).Размышляя о колебаниях жгутиков
,Подвижность клеток и цитоскелет
,2009
, vol.66
(стр.425
—436
),.Характер изгиба хламидомонады жгутиков: IV. Мутанты с дефектами во внутреннем и внешнем плечах динеина указывают на различия в функции плеча динеина
,Cell Motile and the Cytoskeleton
,1987
, vol.8
(стр.68
—75
),,.Анализ движения жгутиков Chlamydomonas : Функция радиально-спицевой системы выявляется путем сравнения жгутиков дикого типа и мутантных
,Journal of Cell Biology
,1982
, vol.92
(стр.722
—732
),.Трехмерный структурный анализ жгутиков / ресничек с помощью криоэлектронной томографии
,Методы Enzymol
,2013
, vol.524
(стр.305
—323
),,,,.Полярность и асимметрия в расположении динеина и родственных структур в аксонеме Chlamydomonas
,Journal of Cell Biology
,2012
, vol.198
(стр.913
—925
),,,,,,.Консервативные структурные мотивы в центральном парном комплексе жгутиков эукариот
,Цитоскелет
,2013
, т.70
(стр.101
—120
),,,,,,.Регуляторная легкая цепь динеина внешнего плеча ресничек у Tetrahymena thermophila
,Journal of Biological Chemistry
,2001
, vol.276
(стр.20048
—20054
),,,.CSC необходим для полной сборки радиальных спиц и подвижности ресничек дикого типа
,Molecular Biology of the Cell
,2011
, vol.22
(стр.2520
—2531
),.Исследование тонкой структуры мерцательного эпителия
,Journal of Morphology
,1954
, vol.94
(стр.221
—281
),.Движение жгутиков и активность аденозинтрифосфатазы в сперме морского ежа, экстрагированной тритоном X-100
,Journal of Cell Biology
,1972
, vol.54
(стр.75
—97
),.Влияние частичной экстракции динеиновых ветвей на движение реактивированных сперматозоидов морского ежа
,Journal of Cell Science
,1973
, vol.13
(стр.337
—357
).Взаимосвязь между тонкой структурой и направлением биений жаберных ресничек пластинчатого моллюска
,Журнал биофизической и биохимической цитологии
,1961
, vol.11
(стр.179
—205
),.О строении жгутиков у некоторых жгутиконосцев
,Журнал биофизической и биохимической цитологии
,1960
, vol.7
(стр.697
—716
),.Динеин: белок с активностью аденозинтрифосфатазы из ресничек
,Science
,1965
, vol.149
(стр.424
—426
),.Субструктура внешнего плеча динеина
,Journal of Cell Biology
,1982
, vol.95
(стр.798
—815
). ,Цилиарное движение
,1928
Cambridge University Press
,.Механизм индуцированного колебаниями жгутика переключения активности динеина в обработанных эластазой аксонемах сперматозоидов морского ежа
,Journal of Cell Science
,2008
, vol.121
(стр.2833
—2843
),,.Вызвание временного биения парализованных жгутиков мутантов Chlamydomonas путем приложения внешней силы
,Подвижность клеток и цитоскелет
,1997
, vol.37
(стр.232
—239
),,,,.Регуляторный комплекс динеина представляет собой нексиновое звено и главный регуляторный узел в ресничках и жгутиках
,Journal of Cell Biology
,2009
, vol.187
(стр.921
—933
),,,,.CSC соединяет три основных комплекса аксонем, участвующих в регуляции динеина
,Molecular Biology of the Cell
,2012a
, vol.23
(стр.3143
—3155
),,,,,,.Криоэлектронная томография выявляет дуплет-специфические структуры и уникальные взаимодействия в динеине I1
,Proceedings of the National Academy of Science
,2012b
, vol.109
(стр.E2067
—E2076
). .Гидродинамические аспекты движения ресничек и жгутиков
,Реснички и жгутики
,1974
Academic Press
(стр.143
—175
),.Физическая модель скольжения микротрубочек в аксонемах ресничек
,Biophysical Journal
,1990
, vol.58
(стр.905
—917
),,.Супрессорные мутации в Chlamydomonas раскрывают регуляторный механизм функции жгутиков
,Cell
,1982
, vol.28
(стр.115
—124
),.Изолированный жгутиковый аппарат Chlamydomonas : характеристика плавания вперед и изменения формы волны и обращения движения ионами кальция in vitro
,Journal of Cell Science
,1978
, vol.33
(стр.235
—253
).Функциональное разнообразие аксонемных динеинов, изученное на мутантах Chlamydomonas
,International Review of Cytology
,2002
, vol.219
(стр.115
—155
),.Субмикромолярные уровни кальция контролируют баланс биений между двумя жгутиками в демембранных моделях Chlamydomonas
,Journal of Cell Biology
,1984
, vol.98
(стр.97
—107
),,.Два типа жгутиковых мутантов Chlamydomonas , в которых отсутствуют различные компоненты динеина внутреннего плеча
,Journal of Cell Biology
,1991
, vol.112
(стр.441
—447
).Большие шаги к пониманию динеина
,Journal of Cell Biology
,2013
, vol.202
(стр.15
—23
).Определение механического состояния аксонемы и интеграция передачи сигналов Ca 2+ с помощью динеина внешнего плеча
,Cytoskeleton
,2010
, vol.67
(стр.207
—213
),. .Аксонемные динеины: сборка, структура и формирование силы
,Справочник по хламидомонаде: подвижность и поведение клеток
,2009
, vol.3
Academic Press
(стр.131
—208
),,,,.Механические свойства внутреннего плеча динеина-f (динеина I1) изучены с помощью тестов подвижности in vitro
,Biophysical Journal
,2007
, vol.93
(стр.886
—894
),,,,.Полиглутамилирование тубулина регулирует подвижность аксонем путем модуляции активности динеинов внутренней части руки
,Current Biology
,2010
, vol.20
(стр.441
—445
),.Chlamydomonas reinhardtii Hydin — это белок центральной пары, необходимый для подвижности жгутиков
,Journal of Cell Biology
,2007
, vol.176
(стр.473
—482
),,,,.Мутации в Hydin нарушают подвижность ресничек у мышей
,Journal of Cell Biology
,2008
, vol.180
(стр.633
—643
),,,,.Структурный механизм динеинового силового удара
,Nature Cell Biology
,2014
, vol.16
(стр.479
—485
).Экспериментальные доказательства гипотезы геометрического сцепления
,Современные темы биологии развития
,2011
, vol.95
(стр.1
—31
),.Свидетельства искривления аксонемы во время биения жгутиков Chlamydomonas
,Подвижность клеток и цитоскелет
,2007
, vol.64
(стр.580
—589
),.Биение жгутиков и ресничек: доказанное и возможное
,Journal of Cell Science
,2010
, vol.123
(стр.519
—528
).Число фибрилл в ресничках зеленых водорослей
,Nature
,1953
, об.171
(стр.485
—486
),.Распространение изгиба приводит к вращению центральной пары у Chlamydomonas reinhardtii жгутиков
,Journal of Cell Biology
,2004
, vol.166
(стр.709
—715
),.Эффекты наложенного изгиба на скольжение микротрубочек в жгутиках сперматозоидов
,Current Biology
,2004
, vol.14
(стр.2113
—2118
).Криоэлектронная томография для изучения организации аксонем
,Методы клеточной биологии
,2009
, vol.91
(стр.1
—39
),.Новый метод структурной маркировки с использованием криоэлектронной томографии и системы биотин-стрептавидин
,Journal of Structural Biology
,2013
, vol.183
(стр.305
—311
),,,.Идентификация внешнего-внутреннего линкера динеина как хаб-контроллера для активности аксонемального динеина
,Current Biology
,2013
, vol.23
(стр.656
—664
),,,.Передача механосигналов между центральным аппаратом и радиальными спицами контролирует активность аксонемального динеина
,Journal of Cell Biology
,2014
, vol.204
(стр.807
—819
),,,,,.Криоэлектронная томография радиальных спиц ресничек и жгутиков
,Journal of Cell Biology
,2011
, vol.195
(стр.673
—687
),,,.Мутации в «динеиновом регуляторном комплексе» изменяют АТФ-нечувствительные сайты связывания динеинов внутреннего плеча в Chlamydomonas аксонемах
,Journal Cell Biology
,1994
, vol.125
(стр.1109
—1117
). .Подвижность жгутиков и регуляторный комплекс динеина
,Динеины: структура
,2012
Биология и болезнь
(стр.337
—365
),,,,.Функции и механизмы моторных белков динеина
,Nature Reviews Molecular Cell Biology
,2013
, vol.14
(стр.713
—726
),.Структура и формирование ресничек и нитей простейших рубца
,Journal of Cell Biology
,1964
, vol.20
(стр.249
—270
),.Молекулярная организация и механизм генерации силы динеина
,FEBS Journal
,2011
, vol.278
(стр.2964
—2979
),.Распространенные Tetrahymena реснички. Система для анализа расположения скользящих и аксонемных спиц
,Journal of Cell Biology
,1976
, vol.71
(стр.589
—605
),.Направление активного скольжения микротрубочек в Tetrahymena ресничках
,Proceedings of the National Academy of Science
,1977
, vol.74
(стр.2045
—2049
).Исследования ресничек: фиксация метахрональной волны
,Journal of Cell Biology
,1963
, vol.18
(стр.345
—365
).Исследования ресничек: II. Исследование дистальной области ствола ресничек и роль нитей в подвижности
,Journal of Cell Biology
,1965
, vol.26
(стр.805
—834
).Морфологические аспекты перистальтики ресничек
,Журнал общей физиологии
,1967
, vol.50
доп.
(стр.241
—258
).Исследования ресничек: 3. Дальнейшие исследования кончика ресничек и модели подвижности ресничек «скользящего филамента»
,Journal of Cell Biology
,1968
, vol.39
(стр.77
—94
),.Расщепление аксонемы ресничек: значение для модели «точки переключения» активности динеинового плеча при движении ресничек
,Подвижность клеток и цитоскелет
,1989
, vol.14
(стр.345
—358
),,.Локальная реактивация жгутиков, экстрагированных тритоном, путем ионтофоретического применения АТФ
,Nature
,1977
, vol.265
(стр.269
—270
),,.Дискретное моделирование ресничек с распределением сингулярности: приложение к эмбриональному узлу и жидкости на поверхности дыхательных путей
,Бюллетень математической биологии
,2007
, vol.69
(стр.1477
—1510
).Hydin seek: обнаружение функции перистальтики ресничек
,Journal of Cell Biology
,2007
, vol.176
(стр.403
—404
),.Радиальные спицы и центральный аппарат: механохимические преобразователи, регулирующие подвижность жгутиков
,Подвижность клеток и цитоскелет
,2004
, vol.57
(стр.8
—17
),.Аденозинтрифосфат-индуцированное скольжение канальцев в обработанных трипсином жгутиках сперматозоидов морского ежа
,Proceedings of the National Academy of Science
,1971
, vol.68
(стр.3092
—3096
),.Вращение и транслокация микротрубочек in vitro , индуцированная динеинами из Tetrahymena ресничек
,Cell
,1988
, vol.52
(стр.459
—469
),.Смещения микротрубочек на концах живых жгутиков
,Подвижность клеток и цитоскелет
,2002
, vol.52
(стр.151
—160
),.Структурная основа образования изгиба ресничек: изменения положения радиальных спиц, сопровождающие скольжение микротрубочек
,Journal of Cell Biology
,1974
, vol.63
(стр.35
—63
),,,,,.Регуляция подвижности ресничек: консервативные протеинкиназы и фосфатазы нацелены и закреплены в аксонеме ресничек
,Архив биохимии и биофизики
,2011
, vol.510
(стр.93
—100
),,.Chlamydomonas жгутиковых мутантов без радиальных спиц и центральных канальцев: структура, состав и функции специфических компонентов аксонемы
,Journal of Cell Biology
,1978
, vol.76
(стр.729
—747
),,,,,,.Аксонемный динеин, особенно важный для движения жгутиков при высокой вязкости: последствия от нового мутанта Chlamydomonas , дефицитного по гену тяжелой цепи динеина DHC9
,Journal of Biological Chemistry
,2005
, vol.280
(стр.41412
—41420
),,,,,,,,,.Комплекс MIA представляет собой консервативный и новый регулятор динеина, необходимый для нормальной подвижности ресничек
,Journal of Cell Biology
,2013
, vol.201
(стр.263
—278
) © Автор (ы) 2014. Опубликовано Oxford University Press от имени Американского института биологических наук. Все права защищены. Для получения разрешений, пожалуйста, пишите на электронную почту: журналы[email protected].Произошла ошибка при установке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файле cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта.